МЯДЕЛЕЦ ОБЩ.ГИСТ.. Литература для медицинских вузов все факультеты о. Д. Мяделец основы цитологии, эмбриологии и общей гистологии
 Скачать 4.75 Mb. Скачать 4.75 Mb.
|

|
Глава 5 ЭМБРИОЛОГИЯ ЧЕЛОВЕКА (МЕДИЦИНСКАЯ ЭМБРИОЛОГИЯ). ПРЕДМЕТ И ЗАДАЧИ МЕДИЦИНСКОЙ ЭМБРИОЛОГИИ. ЗАКОНОМЕРНОСТИ РАННЕГО ЭМБРИОГЕНЕЗА ЧЕЛОВЕКА Эмбриология — наука об эмбриональном развитии организма как части онтогенеза человека. Термин "эмбриология" происходит от греческого "em brio — в оболочках". Медицинская эмбриология изучает закономерности эмбрионального развития человека, причины его нарушений и развития уродств, влияние факторов внешней среды на эмбриогенез, а также механизмы регуляции эмбриогенеза. В последнее время медицинская эмбриология исследует также возможности экстракорпорального оплодотворения и развития зародыша как методов борьбы с бесплодием. Медицинская эмбриология исследует не только морфогенез в описательном его варианте, но также и развитие функций организма и его частей, биохимизм процесса развития и т.д. Поэтому она включает ряд научных направлений: 1. Описательная эмбриология использует методы простого наблюдения и описания процессов нормального развития зародыша. 2. Эволюционная эмбриология изучает закономерности развития организмов в эволюционном плане с целью выяснения общих закономерностей фило- и онтогенеза и применения полученных сведений к процессу развития зародыша человека. 3. Экспериментальная эмбриология использует различные экспериментальные методы для понимания эмбриогенеза человека. 4. Биохимическая эмбриология — направление, изучающее биохимические аспекты развития зародыша, химические факторы регуляции эмбриогенеза и др. 5. Гистологическая эмбриология — направление, целью которого является изучение морфологических аспектов развития зародыша на тканевом, клеточном и субклеточном уровнях. 6. Патологическая эмбриология медико-биологическое направление, в идачу которого входят изучение этиологии, патогенеза и профилактики пороков развития зародыша человека.Этот раздел эмбриологии тесно связан с тератологией - наукой об уродствах, т.к. механизм возникновения уродств может быть понят только на основе закономерностей эмбриогенеза. Задачи эмбриологии человека вытекают из ее предмета и состоят в изучении общих закономерностей развития зародыша человека для того, чтобы обеспечивать нормальное протекание беременности и родов, предотвращать и лечить врожденные уродства. Особое значение эмбриология имеет для такой врачебной специальности, как акушерство. Эмбриональный период как часть онтогенеза включает время от момента оплодотворения яйцеклетки сперматозоидом и образования зиготы до рождения. Весь эмбриогенез подразделяется на 3 периода: начальный (1-я неделя развития), зародышевый (2-8-я недели) и плодный, продолжающийся с 9-й недели внутриутробного развития до рождения ребенка. Это акушерская периодизация эмбриогенеза. Существует также иная периодизация, основанная на учете основных процессов, происходящих в эмбриогенезе - эмбриологическая периодизация). 1. Стадия оплодотворения, в ходе которой образуется одноклеточный зародыш зигота. 2. Стадия дробления. Во время этой стадии образуется многоклеточный зародыш - бластула (бластоциста). 3. Стадия гаструля-ции, в ходе которой образуются зародышевые листки и зародыш, называемый гаструлой, приобретает многослойное строение. 4. Стадия нотогенеза, или образования из зародышевых листков осевого комплекса зачатков (эту стадию выделяют не все исследователи). 5. Стадия гистогенеза, органогенеза и системогенеза. На протяжении этой стадии из тканевых зачатков образуются ткани и органы, идет формирование систем организма. Первая и вторая стадии относятся к начальному периоду эмбриогенеза, зародышевый период включает гаструляцию, нотогенез, гистогенез и органогенез. В плодный период продолжаются процессы гистогенеза и, главным образом, органогенеза и системогенеза. Многие эмбриологи относят к эмбриогенезу также процесс образования половых клеток - проге-нез. Это оправдано, поскольку несмотря на то, что образование половых клеток происходит в организме родителей, нормальный, полноценный прогенез является важнейшим условием развития нового организма. ОСОБЕННОСТИ ЭМБРИОГЕНЕЗА ЧЕЛОВЕКА Эмбриональное развитие млекопитающих и человека характеризуется теми же общими чертами, что и развитие других позвоночных, которые студентам известны из курса биологии. Вместе с тем, находясь на высшей ступени развития животного мира, млекопитающие в целом и человек в особенности характеризуются и специфическими чертами эмбриогенеза, присущими только им. Рассмотрим особенности эмбриогенеза человека. 1. Наиболее важная особенность развития млекопитающих и человека заключается в том, что их развитие происходит внутриутробно при теснейшем взаимодействии организма зародыша и плода с материнским организмом. У приматов эта связь наиболее совершенная. 2. Большая продолжительность эмбрионального периода. У человека он является одним из наиболее продолжительных из всех млекопитающих и длится 280 дней (9 лунных, 10 акушерских месяцев, или 40 недель). 3. Развитие половых клеток в женском организме происходит в эмбриональном периоде. Яйцеклетки в связи с внутриутробным развитием млекопитающих не содержат больших запасов питательных веществ и являются вторично олигоизолецитальными: небольшое количество желтка в них распределено равномерно по ооплазме. Зрелые женские половые клетки выделяются из яичника примерно один раз в месяц в середине менструального цикла. Мужские половые клетки вырабатываются в семенниках постоянно. 4. Осеменение внутреннее, полиспермное. Оплодотворение является моноспермным: свой геном в яйцеклетку при оплодотворении вносит только один сперматозоид. Оплодотворение происходит в яйцеводах и длится несколько часов. 5. Дробление полное, неравномерное, асинхронное. 6. На ранних этапах эмбриогенеза устанавливается тесная связь зародыша с организмом матери. Это происходит во время имплантации (внедрения зародыша в слизистую оболочки матки) и плацентации (образования плаценты как связующего звена между организмом матери и плода). Имплантация у человека, в отличие от других млекопитающих, не поверхностная, а глубокая, интерстициальная (зародыш проникает глубоко в эндометрий, при этом происходит разрушение кровеносных сосудов эндометрия с тем, чтобы стали доступными питательные вещества, содержащиеся в крови матери). В результате у человека в ходе раннего развития зародыша происходит двойная смена типов питания от аутотрофного (за счет потребления питательных запасов зиготы) через гистотрофное (использование секрета эпителия яйцеводов, желез матки, продуктов распада тканей) к гемотрофному. 7. Гаструляция осуществляется в два этапа путем деламинации, иммиграции и частичной инвагинации клеток. 8. Гистогенез и органогенез начинается с 17—20-х суток развития и включает пресомитную, сомитную стадии и стадию дефинитивного гисто-органогенеза. 9. Эмбриогенез млекопитающих и человека характеризуется ранним развитием провизорных органов, характерных для других позвоночных и появлением таких новых провизорных органов, как хорион, плацента, пупочный канатик. Плацента у приматов относится к типу дискоидальных гемохориальных плацент. 10. Весь эмбриогенез человека подразделяется на начальный, зародышевый и плодный периоды, которые характеризуются своими особенностями. 11. Период гистогенеза и органогенеза у человека очень продолжительный, занимает большую часть эмбриогенеза, однако полностью не завершается при рождении ребенка. В связи с этим новорожденные дети в целом являются относительно менее развитыми и более беспомощными, чем родившееся потомство некоторых других млекопитающих. 12. Для эмбрионального развития человека характерно бурное развитие головного мозга, что приводит к высокому индексу цефализации (отношение массы головного мозга к массе плода). МОРФОФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА ПОЛОВЫХ КЛЕТОК. СПЕРМАТОГЕНЕЗ И ОВОГЕНЕЗ. СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА Половыми клетками являются мужская половая клетка сперматозоид и женская половая клетка — яйцеклетка. Половые клетки гаплоидны. СПЕРМАТОЗОИД. Состоит из головки и хвостика и имеет длину 60—70 мкм. В свою очередь, в хвостике различают связующий, промежуточный, главный и дистальный отделы. На обычных гистологических препаратах ядра сперматозоидов обладают резко выраженной базофилией и имеют вытянутую грушевидную форму (рис. 5.1). Ядро характеризуется плотным расположением хроматина, в котором наряду с гистоновыми находятся также простые негистоновые белки, обладающие положительным зарядом. ДНК в хроматине упакована параллельно, поэтому ядро имеет кристаллоподобпую структуру, его объем сведен до минимума. Ядро сперматозода гаплоидно, содержит 22 аутосомы и 1 половую хромосому, которая может быть либо Х-, либо Y-хромосомой. Количество сперматозоидов с Х- или Y-хромосомой примерно одинаковое. Ядерная оболочка сперматозоидов полностью лишена ядерных пор. В передней части ядра под цитолеммой сперматозоида находится ак-росома — производное комплекса Гольджи и аналог лизосомы. Ее мембрана спереди прилежит к цитолемме, а сзади — к ядерной мембране. В акро-соме содержатся ферменты (10—12 различных ферментов), расщепляющие компоненты прозрачной оболочки яйцеклетки: гиалуронидаза, протеазы, гликозидаза, липаза, нейраминидаза, фосфатазы и др. Цитоплазма сперматозоида редуцирована до минимума и очень тонким слоем покрывает ядро. Связующий отдел хвостика содержит проксимальную центриоль. которая прилежит к ядру, располагаясь в углублении ядерной оболочки. Здесь же находится дис-тальная центриоль. От нее отходи! осевая нить — аксонема, имеющая структуру реснички и состоящая ил 9 периферических дуплетов микротрубочек и двух расположенных в центре одиночных микротрубочек Аксонема продолжается во все отделы хвостика, редуцируясь в дис-талыюм отделе. Снаружи напротив каждого дуплета дистальной цент-риоли в связующем отделе находится одна так называемая сегментированная колонна (следовательно, всего количество сегментированных колони равно 9).  В промежуточном отделе хвостика сегментированные колонны продолжаются в 9 плотных волокон. В этом же отделе вокруг аксонемы и плотных волокон в виде спирали располагаются митохондрии. В главной части хвостика два из девяти плотных волокон, расположенные напротив друг друга, резко утолщаются и превращаются в продольные столбы, которые соединяются между собой боковыми ребрами. Формируется внешняя волокнистая оболочка, придающая жесткость и упругость хвостику. Таким образом, сегментированные колонны, плотные волокна, продольные столбы и соединяющие их ребра, а также внешняя волокнистая оболочка образуют каркас хвостика. В дистальном отделе хвостика количество микротрубочек сильно редуцируется. Снаружи хвостик покрыт цитолеммой. Благодаря движению хвостика сперматозоиды могут двигаться со скоростью 1—5 мм в минуту. Электронномикроскопическое строение сперматозоида отражено на рис. 5.2. В промежуточном отделе хвостика сегментированные колонны продолжаются в 9 плотных волокон. В этом же отделе вокруг аксонемы и плотных волокон в виде спирали располагаются митохондрии. В главной части хвостика два из девяти плотных волокон, расположенные напротив друг друга, резко утолщаются и превращаются в продольные столбы, которые соединяются между собой боковыми ребрами. Формируется внешняя волокнистая оболочка, придающая жесткость и упругость хвостику. Таким образом, сегментированные колонны, плотные волокна, продольные столбы и соединяющие их ребра, а также внешняя волокнистая оболочка образуют каркас хвостика. В дистальном отделе хвостика количество микротрубочек сильно редуцируется. Снаружи хвостик покрыт цитолеммой. Благодаря движению хвостика сперматозоиды могут двигаться со скоростью 1—5 мм в минуту. Электронномикроскопическое строение сперматозоида отражено на рис. 5.2.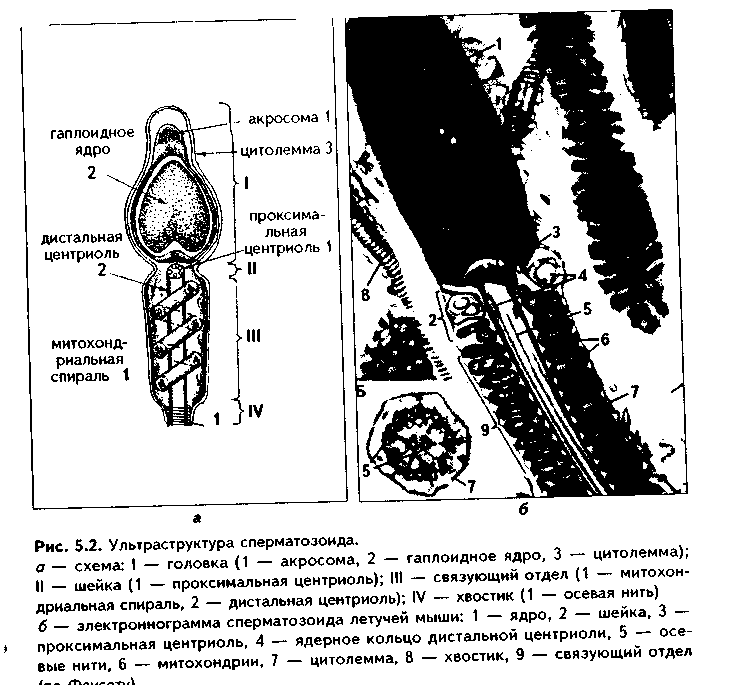 ЯЙЦЕКЛЕТКА. Это женская половая клетка с гаплоидным набором хромосом. В процессе овуляции из яичника выходит овоцит второго порядка с незавершенным мейозом, заблокированным на метафазе 2 мейоти-ческого деления. Этот блок хромосомного аппарата овулировавших овоцитов 2 порядка достаточно стабильный, так что клетки могут длительно сохраняться в метафазе II мейоза. Блокада мейоза снимается лишь при оплодотворении, и завершение мейоза с образованием зрелой яйцеклетки происходит сразу после него. Яйцеклетка (оотида) человека имеет округлую форму (рис. 5.3, 5.4). Ее диаметр равен около 130 мкм. Содержит 23 хромосомы, одна из кото-рых половая Х-хромосома. При электронномикроскопическом исследовании в цитоплазме яп цеклетки выделяются митохонд рии, комплекс Гольджи, хорошо развитые гранулярная и аг-ранулярная эндоплазматическая сеть, а также включения: трофические (гранулы вителлина, липидные) и пигментные. Снаружи яйцеклетка окружена плазматической мембраной. Под ней располагается толстый слой цитоплазмы толщиной 2— 3 мкм. Вместе с цитолеммой его называют кортикальным слоем. В кортикальном слое находятся кортикальные гранулы, содержащие различные ферменты, в том числе и овопероксидазу, действие которой на блестящую оболочку после оплодотворения резко изменяет свойства последней. Кортикальный слой играет важную роль в организации яйцеклетки, а также в оплодотворении. 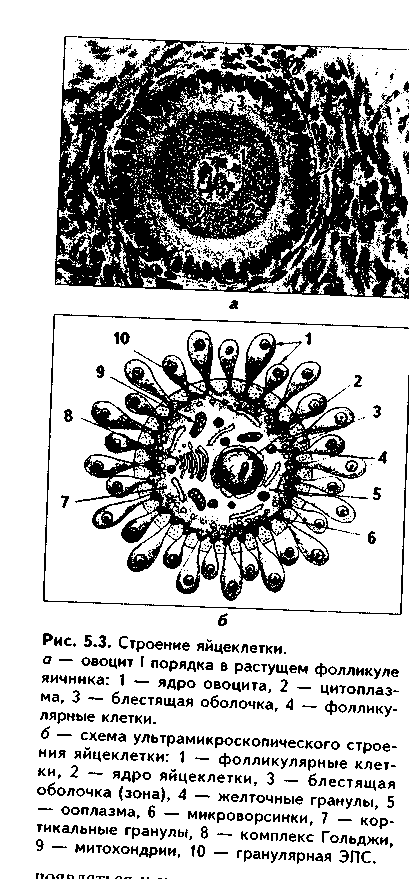 Яйцеклетки имеют хорошо развитый и своеобразно организованный цитоскелет. Его компоненты связаны с цитолеммой и вызывают постоянную модификацию поверхности клетки, в которой могут появляться и исчезать микроворсинки, меняться локализация рецепторов. Яйцеклетки окружены блестящей оболочкой и слоем фолликулярных клеток. Между блестящей зоной и цитолеммой овоцита имеется небольшое перивителлиновое пространство, которое существенно увеличивается после оплодотворения, поскольку в него выделяются содержимое кортикальных гранул. Фолликулярные клетки через отверстия в блестящей оболочке посылают к цитолемме овоцита свои отростки. В ряду позвоночных в зависимости от наличия желтка, его количества и распределения по ооплазме различают алицетальные (без желтка) и леци-тальные (с желтком) яйцеклетки.  Лецитальные клетки делятся на олиголецитальные (маложелтковые) и полилецитальные (многожелтковые). В зависимости от распределения желтка по ооплазме яйцеклетки делятся на изолецитальные, умеренно телолецитальные и резко телолецитальные. В изолецитальных яйцеклетках желток распределен равномерно. В уме ренно телолецитальных яйцеклетках желток находится на одном полюсе, который называется вегетативным. На другом полюсе — анимальном — лс жат органеллы и ядро. В резко телолецитальных клетках вегетативный но люс выражен особенно сильно и занимает подавляющую часть клетки. Яйцеклетки млекопитающих, в том числе и человека, являются олиго-изолецитальными: содержат очень мало желтка, который равномерно рас пределен по ооплазме. РОЛЬ ЯДРА И ЦИТОПЛАЗМЫ В ПЕРЕДАЧЕ НАСЛЕДСТВЕННОЙ ИНФОРМАЦИИ Наследственная информация хранится в ядре половых клеток, в хромосомах. Она закодирована в триплетах ДНК — генах. Ядро половых клеток выполняет три основные функции, присущие ему и в других клетках: 1. Хранение и передача наследственной информации. 2. Реализация наследственной информации. 3. Контроль за синтетическими процессами, осуществляемыми в цитоплазме. Первая функция обеспечивается путем репликации молекул ДНК. Это позволяет при митозе двум дочерним клеткам получать одинаковые объемы генетической информации. Для сохранения генетической информации в неизмененном виде в ядре имеются ферменты, ликвидирующие повреждения молекул ДНК. Генотип половых клеток содержит только половину программы развития, строения и функционирования индивидуума, полная программа создается и реализуется только после оплодотворения. Вторая и третья функции обеспечиваются процессами транскрипции на молекулах ДНК различных информационных, транспортных и рибосо-мальных РНК. В ядре происходит также образование субъединиц рибосом путем соединения синтезированных в ядрышке рибосомных РНК с рибо-сомными белками. Эти белки синтезируются в цитоплазме и переносятся в ядро. Субъединицы рибосом выходят в цитоплазму и соединяются с информационной РНК. После этого происходит синтез белков, необходимых для дифференцировки и специализации, а также выполнения клетками функций. Кроме ядра, некоторое количество ДНК содержится в цитоплазме в митохондриях, которые могут самостоятельно синтезировать определенные белки для собственных нужд. В цитоплазме вырабатываются вещества (триггерные белки), способные проникать в ядро и регулировать активность генома клеток. Таким образом, цитоплазма и ядро тесно взаимосвязаны is выполнении функций по хранению и передаче наследственной информации. Итак, программа эмбрионального развития индивидуума заключена в ДНК зиготы и реализуется на основе экспрессии и репрессии генов, дифференцировки клеток под влиянием зпигеномных стимулов и стимулов микроокружения. ПРОГЕНЕЗ Прогенезом называется процесс образования половых клеток, или га-метогенез. В свою очередь, гаметогенез делится на сперматогенез (образование сперматозоидов) и овогенез (образование яйцеклеток). Развитие половых клеток в эмбриогенезе человека начинается довольно рано. Они возникают во внезародышевой желточной энтодерме в конце 3-й недели эмбриогенеза. Позднее эти клетки (они называются гонобластами) мигрируют в закладку половых желез на медиальной поверхности первичной почки и принимают участие в образовании половых желез — гонад. СПЕРМАТОГЕНЕЗ. Процесс развития мужских половых клеток — сперматозоидов — называется сперматогенезом. В сперматогенезе различают 4 фазы: размножения, роста, созревания и формирования (рис 5.5). На протяжении периода размножения мужские половые клетки представлены сперматогониями. Это мелкие округлые клетки, делящиеся митозом. Они подразделяются на темные и светлые сперматогонии. Темные сперматогонии являются истинными стволовыми клетками, устойчивы к действию вредных факторов и способны совершать редкие митотические деления. Светлые сперматогонии подразделяются на А и В-сперматогонии. А-снерматогонии являются полустволовыми клетками, способными к частым митотическим делениям. При делении каждой такой сперматогонии могут возникать либо две А-сиерматогонии, либо одна А- и одна В-спер-матогопия. В-снерматогонии также способны митотически делиться, но при этом не происходит цитотомии, и клетки оказываются связанными между собой цитоилазматическими мостиками. При этом возникают клоны (ассоциации) клеток. После некоторой паузы В-сперматогонии вступают в период роста, в течение которого превращаются в сперматоциты первого порядка. Для периода роста характерен значительный рост ядра и цитоплазмы развивающихся клеток, их размеры увеличиваются в четыре и более раз. Сперматоциты I порядка митотически не делятся, но вступают в период созревания, который состоит из двух последовательных делений мейоза (мейоз I и мейоз II). 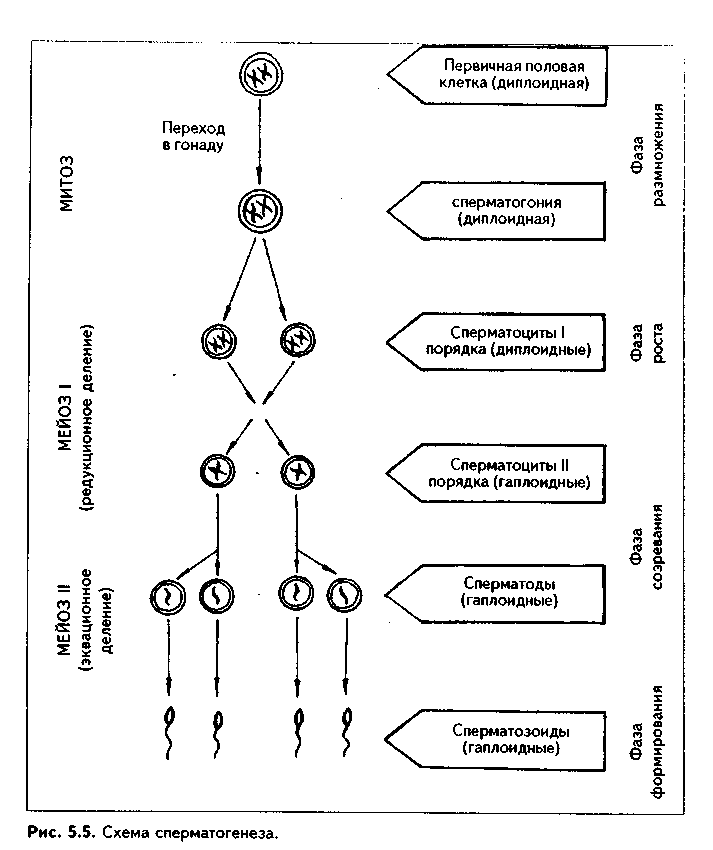 Мейоз I называется редукционным делением, так как в ходе его происходит уменьшение в два раза количества хромосом с формированием гаплоидного генома. Мейоз I имеет сложную профазу, состоящую из 5 стадий: лептотены, зиготены, пахитены, диплотены и диакинеза В стадию лептотены хромосомы спирализуются и становятся видны в виде длинных тонких нитей. В стадию зиготены гомологичные хромосомы тесно сливаются друг с другом. В это время в них происходит кросинговер — обмен генами. В стадию пахитены хромосомы сильно спирализуются и укорачиваются. Диплотена характеризуется расщеплением хромосом на хроматиды и образованием тетрад. В диакинезе хромосомы еще более утолщаются и несколько отходят друг от друга. Таким образом, в профазу мейоза 1 происходит подготовка к редукции числа хромосом. В метафазу гомологичные нары хромосом располагаются в экваторе клетки. Это ключевой момент подготовки к редукции хромосом. В анафазу к полюсам отходят целые гомологичные хромосомы, и этот момент является началом редукции генома. В телофазу происходит цитотомия, в результате чего образуются два сперматоцита II порядка с гаплоидным набором хромосом. Их хромосомы состоят из двух хроматид. Второе деление мейоза (мейоз II) называется эквационным, или уравнительным. Оно начинается сразу после мейоза I и протекает по типу обычного митоза. В анафазу мейоза II к полюсам отходят хроматиды, а в результате телофазы образуются сперматиды, содержащие вместо хромосом хроматиды. Сперматиды, как и сперматоциты второго порядка, содержат гаплоидный набор хромосом, каждая из которых представлена одной хроматидой. Все образующиеся в процессе сперматогенеза клетки (В-сперматого-нии, сперматоциты I и II порядка, а также сперматиды) остаются связанными между собой цитоплазматическими мостиками в клеточные ассоциации, или клоны. Окончательное разделение клеток происходит в фазу формирования. Сохранение цитоплазматических мостиков между клетками имеет очень большой биологический смысл. Оказывается, что для полноценной дифференцировки сперматозоидов необходим весь диплоидный геном и продукты его деятельности. Во-первых, потому, что в исходном диплоидном геноме могут содержаться дефектные, летальные аллели генов, и клетка, получившая их, погибнет, если не будет обеспечена продуктами нормального аллеля, находящегося в ядрах клеток, его получивших. Во-вторых, как известно, одни мужские половые клетки получают Х-, другие — Y- половую хромосому. Каждая из них содержит много важных генов, необходимых для развития сперматозоидов. Поэтому благодаря цитоплаз-матическим мостикам развивающиеся мужские половые клетки получают продукты деятельности диплоидного генома. Фаза формирования является самой продолжительной фазой сперматогенеза. В процессе ее из сперматид образуются сперматозоиды. Часто эту фазу называют спермиогенезом. Она длится дольше всех остальных фаз (около 50 суток). Процесс спермиогенеза начинается с образованием из комплекса Гольджи вначале акробласта, а затем акросомы, которая содержит ферменты для разрушения яйцевых оболочек. Центросома, состоящая из двух центриолей, перемещается в противоположный полюс. Проксимальная центриоль прилежит к ядру, а дистальная делится на две части. Из одной части образуется жгутик, который превращается в осевую нить хвостика. Вторая часть играет роль базального тельца. Образуются эле-менты нитоскелета: сегментир<| ванные колонны, плотные волок на, продольные столбы с ребрами. Цитоплазма сперматозоид.! сильно редуцируется, а ядро ста новится вытянутым, компактным и гипербазофильным. На заключительных этапах формирования сперматозоиды отделяются от соединяющей их друг с другом общей цитоплазмы и становятся свободными. Оставшийся после отделения объем цитоплазмы (остаточные тельца) подвергается фагоцитозу. 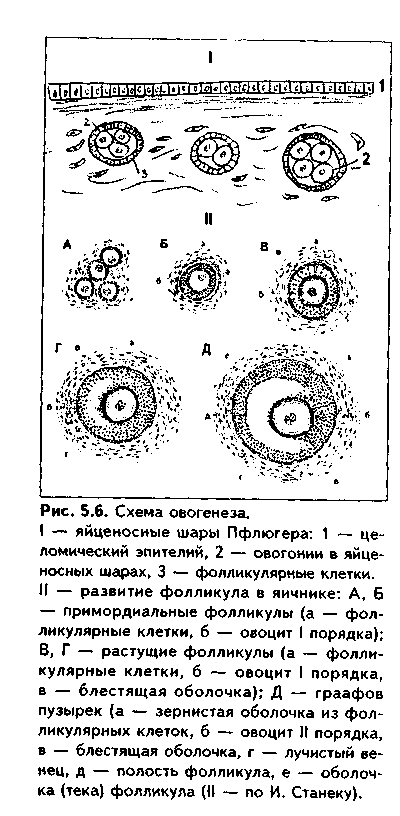 |