МЯДЕЛЕЦ ОБЩ.ГИСТ.. Литература для медицинских вузов все факультеты о. Д. Мяделец основы цитологии, эмбриологии и общей гистологии
 Скачать 4.75 Mb. Скачать 4.75 Mb.
|

|
Глава 3 ЦИТОЛОГИЯ. КЛЕТКА КАК СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ЕДИНИЦА ТКАНИ. ОБЩИЙ ПЛАН СТРОЕНИЯ. ЦИТОПЛАЗМА, СТРОЕНИЕ ОРГАНЕЛЛ И ВКЛЮЧЕНИЙ Цитология — наука о клетке. Она изучает строение и функции тканевых клеток у многоклеточных организмов, одноклеточные организмы, процессы воспроизводства, роста клеток, их регенерации, приспособления к условиям внешней среды и другие процессы, позволяющие судить об общих для всех клеток свойствах и функциях. Клетка — это элементарная структурная единица организма, состоящая из ядра, цитоплазмы и ограниченная клеточной оболочкой, способная выполнять все функции, характерные живому: обмен веществ и энергии, размножение, рост, раздражимость, сократимость, хранение генетической информации и ее передачу. ОСНОВНЫЕ ПОЛОЖЕНИЯ КЛЕТОЧНОЙ ТЕОРИИ И ЕЕ ЗНАЧЕНИЕ Клеточная теория явилась одним из наиболее важных открытий в биологии, перевернувшим существовавшие до нее представления о живой материи. Она дала толчок бурному развитию цитологии, гистологии и эмбриологии и является ее основополагающим учением. Клеточная теория была сформулирована в 1838 году немецкими учеными М. Шлейденом и Т. Шванном, а в дальнейшем развита Вирховым. М. Шлейдеи (1838) создал так называемую теорию цитогенеза, в которой впервые связал возникновение новых клеток не с их оболочкой, а с содержимым и прежде всего с ядром. После этого Т. Шванн (1838) показал, что в явлении цитогенеза скрывается общий принцип развития микроскопических структур всех организмов, позволяющий сделать заключение о принципиальном сходстве клеток всех тканей и органов. Тем самым Т. Шванн обосновал, исходя из генетического принципа, клеточную теорию. Наконец Р. Вирхов в 1859 г. пересмотрел и развил клеточную теорию, выдвинув вместо представлений о цитогенезе положение "всякая клетка из клетки". Однако разработке клеточной теории предшествовали труды многих ученых. В 1824—1827 гг. французские учение А. Дютроше, Ф. Распайль и П. Тюрпен высказали предположение, что мешочки и пузырьки (т.е. клетки) являются элементарными структурными единицами всех растительных и животных тканей. Особо следует отметить чешского ученого Я. Пуркине, который в определенной степени предвосхитил создание клеточной теории. Он в 1837 г. создал теорию "ядросодержащих зернышек", т.е. клеток. Русский гистолог П.Ф. Горянинов на протяжении 1834—1847 гг. сформулировал принцип, согласно которому клетка является универсальной моделью организации живых организмов. В настоящее время главные положения клеточной теории остаются незыблемыми. Однако они существенно дополнены новейшими сведениями о строении клеток, их размножении и гибели, взаимодействии клеток при выполнении своих функций и т.д. Современная клеточная теория включает такие положения: 1. Клетка является наименьшей единицей живого. 2. Клетки разных организмов имеют похожее строение. 3. Размножение клеток происходит путем деления материнской клетки (omnia cellula e cellule — каждая клетка — из клетки). 4. Многоклеточные организмы состоят из сложных ансамблей клеток и их производных. Значение клеточной теории состоит в следующем: 1. Она явилась фундаментом для развития многих биологических дисциплин, прежде всего цитологии, гистологии, эмбриологии, физиологии, а также патологии. 2. Позволила понять механизмы онтогенеза — индивидуального развития организмов. 3. Явилась основой для материалистического понимания жизни, окружающего мира. 4. Явилась основой для объяснения эволюции организмов. СТРОЕНИЕ КЛЕТКИ Клетка может существовать как самостоятельно, так и в составе тканей многоклеточных животных и растений. В составе тканей клетки являются важнейшим тканевым элементом. Все клетки делятся па прокариотические и эукариотические. Прокариота ческие клетки не | имеют ядерной оболочки, не со-*. держат органелл, ядра. Вся гене- Втическая информация у них хранится в замкнутой в кольцо двойной цепи ДНК. Прокариотические клетки окружены жест-£ кой клеточной стенкой. Они ли-I шены митотического аппарата. К прокариотам относятся некоторые бактерии и водоросли. Все остальные клетки являются эукариотическими. Они отличаются от прокариотов наличием хромосом, системы внутриклеточных мембран, из которых построены органеллы. Цитоплазматические мембраны отграничивают также ядро. Имеется митотический аппарат. Организм взрослого человека состоит из примерно 10" клеток, подразделяющихся на более чем 200 типов, существенно различающихся как строением, так и функциями. Однако при имеющихся несомненных различиях клетки всех этих типов имеют общие черты строения. Эукариотическая клетка состоит из таких компонентов (рис. З.1.):
В свою очередь, каждый из этих трех компонентов клетки состоит из нескольких частей. Клеточная оболочка образована трех частями: снаружи располагается гликокаликс, затем идет цитоплазматическая мембрана (цитолемма, плазмолемма), а иод ней находится подмембранный слой опорно-сократительных структур. Цитоплазма также состоит из трех частей: гиалоплазмы, органелл и включений. Ядро построено из четырех компонентов: 1) ядерной оболочки, или кариолеммы, 2) ядрышка, 3) хроматина (хромосом), 4) ядерного сока (кариолимфы). 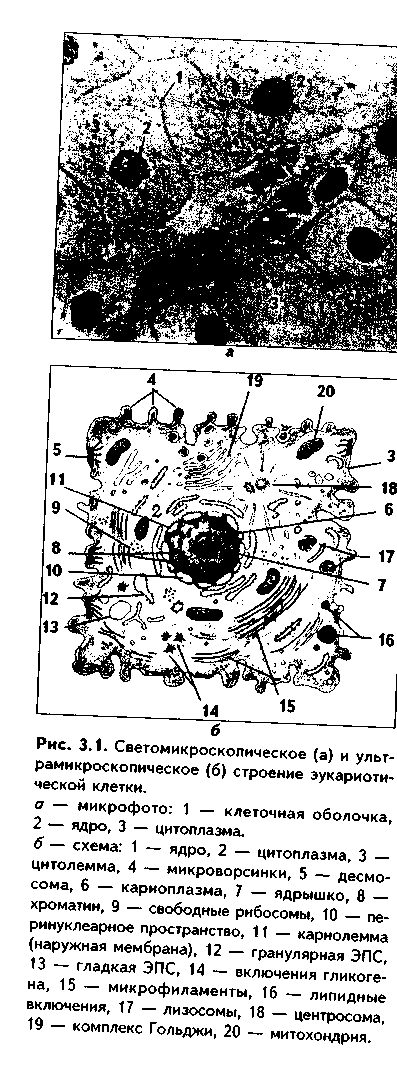 КЛЕТОЧНАЯ ОБОЛОЧКА Основной частью клеточной оболочки является цитоплазматическая мембрана (цитолемма), которая имеет строение элементарной биологической мембраны, являясь самой толстой из всех других клеточных мембран (7,5-11 нм). БИОЛОГИЧЕСКИЕ МЕМБРАНЫ — это липопротеидные образования, которые ограничивают клетку снаружи и формируют некоторые органеллы, а также оболочку ядра. В электронном микроскопе имеют трехслойную структуру (два темных слоя разделены светлым слоем) из-за особого расположения структурных компонентов (рис. 3.2). Основными химическими компонентами клеточных мембран являются липиды (40%), белки (50%) и углеводы (10%). Молекулы липидов мембран состоят из двух частей: гидрофильной и гидрофобной, т.е. полярны. С полярностью липидов мембран связана их проницаемость для веществ. Неполярные соединения легко проникают через нее, тогда как полярные (например, белки) могут проникать в клетку только путем эндоцитоза (см. ниже). В мембранах липиды образуют ли-пидный бислой, в котором молекулы липидов имеют характерное расположение: гидрофобные концы (хвостики) спрятаны внутрь бислоя, а гидрофильные части находятся снаружи. Хвостики липидов образуют центральный светлый слой мембран. Среди липидов (липоидов) мембран выделяют фосфолипиды, сфинголипиды, а также холестерин. Из мембранных фосфолипидов может высвобождаться арахидоновая кислота, являющаяся предшественником ряда биологически активных веществ и гормоноидов: простагландинов, тромбоксанов, лейкотриенов и других, выполняющих множество функций (медиаторы воспаления, вазоактивные факторы, вторичные посредники и др.). Белки мембран разделяются на 3 основных группы: поверхностные белки расположены или снаружи, или изнутри липидного бислоя; они непрочно связаны с поверхностью мембраны и чаще находятся вне липидного бислоя; интегральные (трансмембранные) белки проходят через всю толщину бислоя; полуинтегральные белки проникают только до половины липидного бислоя. По функции белки мембран могут быть белками-ферментами, белками-реценторами, транспортными, а также структурными белками. Белковые молекулы располагаются в липидном бислое мозаично и могут "плавать" в "липидном море" наподобие айсбергов. При межклеточных взаимодействиях может происходить концентрация их на взаимодействующих участках нитолеммы в виде агрегатов (так называемый кэппинг). В перемещении белков важную роль играют элементы цитоскелета (микро-филаменты). Описанная модель строения биологических мембран называется жидкомозаичной квазикристаллической (мембрана имеет кристаллоподобную структуру, в которой, однако, белки не закреплены, а подвижны благодаря текучести мембраны). Углеводы мембран входят в их состав не самостоятельно, а являются частями сложных белков и липидов-гликопротеидов и гликолипидов. ФУНКЦИИ БИОЛОГИЧЕСКИХ МЕМБРАН 1. Разграничительная — отделяют клетку от внеклеточной среды, ядро от цитоплазмы, содержимое органелл от их микросреды и т.д. 2. Барьерно-защитная: защищают внутреннюю среду клетки от действия вредных внешних факторов. 3. Рецепторная (см. рис. 3.2). 4. Транспортная: транспорт веществ в клетку — эндоцитоз, и из клетки — экзоцитоз. 5. Участие в межклеточных взаимодействиях: формирование межклеточных контактов, дистантные взаимодействия между клетками. Второй частью клеточной оболочки является гликокаликс (Рис. 3.2). Он представлен углеводными концами сложных белков (гликопротеинов) и сложных липидов (липопротеинов), входящих в состав цитомембраны. В гликокаликсе располагаются также поверхностные белки мембран, полуинтегральные белки. Их функциональные участки находятся в гликокаликсе. Эти белки могут играть роль ферментов. В гликокаликсе находятся рецепторы гистосовместимости, иммуноглобулины, могут адсорбироваться ферменты, рецепторы гормонов. Функции гликокаликса: 1. Рецепторная (распознавание молекул соседних клеток и межклеточного вещества); 2. Межклеточные (адгезивные) контакты и взаимодействия; 3. Ориентация белков в мембране; 4. Участие в транспорте веществ. Третий компонент клеточной оболочки — подмембранный слой опорно-сократительных структур. В его состав входят сократительные структуры — актиновые филаменты, а также опорный аппарат — кера-тиновые филаменты, микротрубочки. Подмембранный слой тесно связан с цитоскелетом с одной стороны, и рецепторами гликокаликса — с другой. Функции подмембранного слоя: поддержание формы клетки, создание ее упругости, изменения клеточной поверхности, за счет чего клетка участвует в эндо- и экзоцитозе, фагоцитозе, движении, секреции. С другой стороны, подмембранный слой связывает клеточную поверхность с компонентами цитоплазмы, поддерживает их упорядоченное расположение. ПОНЯТИЕ О ЦИТОРЕЦЕПТОРАХ Рецепторы представляют собой белковые молекулы на поверхности клетки, в ее цитоплазме или ядре, которые специфически реагируют с лигандами (гормонами, нейромедиаторами, факторами роста, цитокинами) или другими клетками. В соответствии со своей локализацией рецепторы делятся на поверхностные и внутриклеточные, а внутриклеточные подразделяются на цитоплазматические и ядерные. Поверхностные рецепторы образованы поверхностными белками цито-мембран, а также гликокаликсом. Они предназначаются для полярных лиганд, т.е. веществ, которые не могут проникнуть через клеточную мембрану внутрь клетки и оказывают свое действие на нее через систему внешних рецепторов и вторичных посредников. Подразделяются на каталитические рецепторы, рецепторы, связанные с ионными каналами, рецепторы, связанные с G-белками, и рецепторы, связывающие молекулы внеклеточного матрикса с цитоскелетом. Гликокаликс образует своеобразные "антенны", которые состоят из нескольких моно(олиго)сахаридных участков. Эти участки имеют разную конфигурацию, благодаря чему могут связываться с самыми различными химическими веществами. "Антенны" распознают различные внешние сигналы: молекулы гормонов, нейромедиаторов, факторов роста, цитокинов, генетически чуждые вещества и др. Рецепторные белки и углеводные участки часто связаны с ферментами (каталитические рецепторы). Такие рецепторные белки являются трансмембранными и состоят из рецепторного и каталитического участков. В качестве примера можно привести протеинкиназы (например, тирозин-киназа). Эти ферменты активируют внутриклеточные белки, что ведет к образованию второго посредника (мессенджера), передающего внешние сигналы в клетку, изменяя ее метаболизм, усиливая или ослабляя обмен веществ, синтез секрета. Так построены рецепторы инсулина, факторов роста и др. Мембранные рецепторы могут изменять проницаемость мембран для ионов, что ведет к формированию электрического импульса (рецепторы к нейромедиаторам). Это так называемые рецепторы, связанные с ионными каналами. Рецепторы также контролируют поступление в клетку различных молекул, связывают молекулы внеклеточного матрикса с компонентами цитоске-лета (рецепторы, связывающие молекулы внеклеточного матрикса с цитоскелетом). К таким рецепторам относят, например, интегрины. Они относятся к молекулам адгезии клеток (МАК). Интегрины — трансмембранные белки, воспринимающие молекулы внеклеточного матрикса, в частности, фибронектина и ламинина. В свою очередь, фибронек-тин связывается с другими молекулами внеклеточного матрикса (фибрином, коллагеном, гепарином и др.), а интегрин при помощи ряда других белков — с цитоскелетом. Таким образом, влияние молекул внеклеточного матрикса может передаваться на компоненты цитоскелета. Под влиянием раздражения этого вида рецепторов может изменяться состояние подмембранного слоя, и клетка может начать движение, а также экзоцитоз, эндоцитоз и другие виды деятельности. Особый вид поверхностных рецепторов — рецепторы, связанные с G-белками. Это трансмембранные белки, которые могут быть связаны либо с ионным каналом, либо с ферментом. Состоят из двух частей: рецепторной, взаимодействующей с сигнальными молекулами, и субъединиц G-белка ос, Р, у. G-белки — белки, связывающие гуанозинтрифосфат (ГТФ). После связывания с сигнальной молекулой комплекс G-белков передает сигнал на ассоциированный с цитолеммой фермент аденилатциклазу, которая синтезирует вторичный посредник циклический аденозинмонофосфат (цАМФ). В качестве вторичного посредника могут выступать и молекулы кальция. Через рецепторы, связанные G-белками, опосредуется действие на клетку подавляющего большинства гормонов и нейромедиаторов. Внутриклеточные рецепторы находятся внутри клетки — в гиалоплазме, на мембранах органелл (цитоплазматические рецепторы), в ядре (ядерные рецепторы). Они предназначены для гормонов и других биологически активных веществ, которые в силу неполярности своих молекул могут легко проникать внутрь клетки (стероидные и тиреоидные гормоны и др.). Особый интерес представляют ядерные рецепторы. С этими рецепторами связываются такие гормоны, как стероидные, тиреоидные, витамин D,. Молекулы таких рецепторов состоят из 2 участков: участок для связывания с гормоном и участок, взаимодействующий со специфическими участками ДНК в ядре. Ядерные рецепторы являются факторами транскрипции. Некоторые из них относятся к протоонкогенам — генам нормального генома, регулирующим пролиферацию клеток органов-мишеией, их дифференцировку и межклеточные взаимодействия. В результате соматических мутаций в протоонкогенах может происходить злокачественное перерождение клеток. Внутриклеточные рецепторы могут находиться также на мембранах органелл, например, на митохондриях содержатся рецепторы к тиреоидным гормонам и т.д. 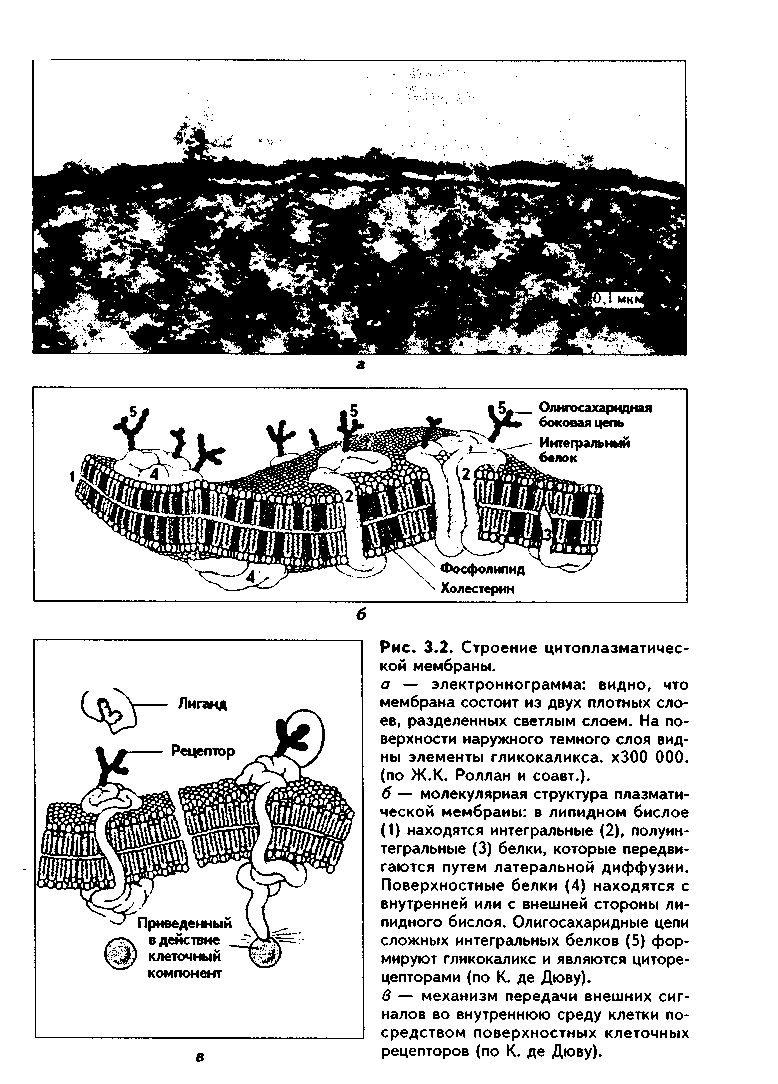 МОЛЕКУЛЫ АДГЕЗИИ КЛЕТОК (МАК) С деятельностью поверхностных рецепторов клеток связан такой феномен, как клеточная адгезия. Адгезия — процесс взаимодействия специфических гликопротеинов соприкасающихся плазматических мембран распознающих друг друга клеток или клеток и внеклеточного матрикса. В том случае, если гликоиротеины при этом образуют связи, происходит адгезия, а затем формирование прочных межклеточных контактов или контактов клетки и межклеточного матрикса. Все молекулы клеточной адгезии подразделяются на 5 классов. 1. Кадгерины. Это трансмембранные гликопротеины, использующие для адгезии ионы кальция. Отвечают за организацию цитоскелета, взаимодействие клеток с другими клетками. 2. Интегрины. Как уже отмечалось, интегрины представляют собой мембранные рецепторы для белковых молекул внеклеточного матрикса — фибронектина, ламинина и др. Связывают внеклеточный матрикс с цитос-келетом при помощи внутриклеточных белков талина, винкулина, а-акти-нина. Функционируют как клеточно-виеклеточные, так и межклеточные адгезионные молекулы. 3. Селектины. Обеспечивают прилипание лейкоцитов к эндотелию сосудов и тем самым — лейкоцитарно-эндотелиальпые взаимодействия, миграцию лейкоцитов через стенки сосудов в ткани. 4. Семейство иммуноглобулинов. Эти молекулы играют важную роль в иммунном ответе, а также в эмбриогенезе, заживлении ран и др. 5. Гоминговые молекулы. Обеспечивают взаимодействие лимфоцитов с эндотелием, их миграцию и заселение специфических зон иммунокомпс-тептных органов. Таким образом, адгезия является важным звеном клеточной рецепции, играет большую роль в межклеточных взаимодействиях и взаимодействиях клеток с внеклеточным матриксом. Адгезионные процессы абсолютно необходимы при таких общебиологических процессах, как эмбриогенез, иммунный ответ, рост, регенерация и др. Они участвуют также в регуляции внутриклеточного и тканевого гомеостаза. ЦИТОПЛАЗМА ГИАЛОПЛАЗМА. Гиалоплазму называют также клеточным соком, цитозолем, или клеточным матриксом. Это основная часть цитоплазмы, составляющая около 55% объема клетки. В ней осуществляются основные клеточные обменные процессы. Гиалонлазма является сложной коллоидной системой и состоит из гомогенного мелкозернистого вещества с низ-кой электронной плотностью. Она состоит из воды, белков, нуклеиновых кислот, полисахаридов, липидов, неорганических веществ. Гиалоплазма может менять свое агрегатное состояние: переходить из состояния жидкого (золь) в более плотное — гель. При этом может изменяться форма клетки, ее подвижность и обмен веществ. Функции гиалонлазмы: 1. Метаболическая — метаболизм жиров, белков, углеводов. 2. Формирование жидкой микросреды (матрикса клетки). 3. Участие в движении клетки, обмене веществ и энергии. ОРГАНЕЛЛЫ. Органеллы — это второй важнейший обязательный компонент клетки. Важным признаком органелл является то, что они имеют постоянное строго определенное строение и функции. По функциональному признаку все органеллы делятся на 2 группы: |