МЯДЕЛЕЦ ОБЩ.ГИСТ.. Литература для медицинских вузов все факультеты о. Д. Мяделец основы цитологии, эмбриологии и общей гистологии
 Скачать 4.75 Mb. Скачать 4.75 Mb.
|

|
1. Органеллы общего значения. Содержатся во всех клетках, поскольку необходимы для их жизнедеятельности. Такими органеллами являются: митохондрии, эндоплазматическая сеть (ЭПС) двух видов, комплекс Голь-джи (КГ), центриоли, рибосомы, лизосомы, пероксисомы, микротрубочки и микрофиламенты. 2. Органеллы специального значения. Есть только в тех клетках, которые выполняют специальные функции. Такими органеллами являются миофибриллы в мышечных волокнах и клетках, нейрофибриллы в нейронах, жгутики и реснички. По структурному признаку все органеллы делятся на: 1) органеллы мембранного типа и 2) органеллы немембранного типа. Кроме того, немембранные органеллы могут быть построены по фибриллярному и гранулярному принципу. В органеллах мембранного типа основным компонентом являются внутриклеточные мембраны. К таким органеллам относятся митохондрии, ЭПС, КГ, лизосомы, пероксисомы. К немембранным органеллам фибриллярного типа относятся микротрубочки, микрофиламенты, реснички, жгутики, центриоли. К немембранным гранулярным органеллам относят рибосомы, полисомы. МЕМБРАННЫЕ ОРГАНЕЛЛЫ ЭНДОПЛАЗМАТИЧЕСКАЯ СЕТЬ (ЭПС) - мембранная органелла, описанная в 1945 году К. Портером. Ее описание стало возможно благодаря электронному микроскопу. ЭПС — это система мелких каналов, вакуолей, мешочков, образующих в клетке непрерывную сложную сеть, элементы которой часто могут формировать кажущиеся на ультратонких срезах изолированными вакуоли. ЭПС построена из мембран, более тонких, чем цитолемма, и содержащих больше белка из-за находящихся в ней многочисленных ферментных систем. Есть 2 вида ЭПС: гранулярная (шероховатая) и агранулярная, или гладкая. Оба вида ЭПС могут взаимно переходить друг в друга и функционально связаны между собой так называемой переходной, или транзиторной, зоной. Гранулярная ЭПС (рис. 3.3) содержит на своей поверхности рибосомы (полисомы) и является органеллой биосинтеза белка. Полисомы или рибосомы связываются с ЭПС при помощи так называемого причального белка (docking protein). При этом в мембране ЭПС имеются специальные интегральные белки рибофорины, также связывающие рибосомы и формирующие гидрофобные трапемембранные каналы для транспорта синтезированной полипентидной цени в просвет гранулярной ЭПС. Гранулярная ЭПС видна только в электронном микроскопе. В световом микроскопе признаком развитой гранулярной ЭПС служит базофилия цитоплазмы. Гранулярная ЭПС имеется в каждой клетке, но степень ее развития различна. Она максимально развита в клетках, синтезирующих белок на экспорт, т.е. в секреторных клетках. Максимального развития гранулярная ЭПС достигает в нейроцитах, в которых ее цистерны приобретают упорядоченное расположение. В этом случае на светомикроскопическом уровне она выявляется в виде закономерно расположенных участков базофилии цитоплазмы, называемых базофилыюй субстанцией Ниссля. 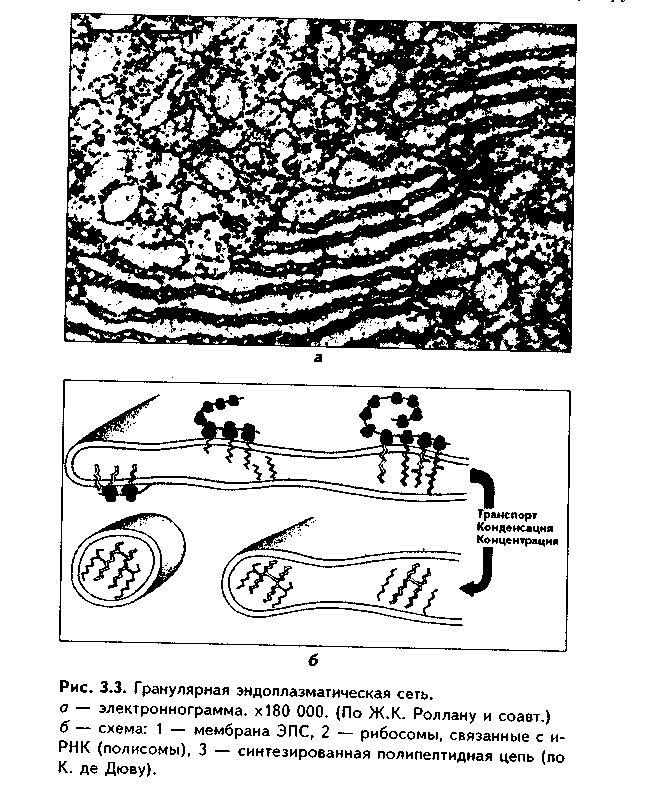 Функция гранулярной ЭПС — синтез белка на экспорт. Кроме того, в ней происходят начальные посттрансляционные изменения полипептидной цепочки: гидроксилирование, сульфатирование и фосфорилиро-вание, гликозилирование. Последняя реакция особенно важна, т.к. приводит к образованию гликопротеинов — наиболее частого продукта клеточной секреции. Агранулярная (гладкая) ЭПС представляет собой трехмерную сеть канальцев, не содержащих рибосомы. Гранулярная ЭПС может без перерыва переходить в гладкую ЭПС, но может существовать как самостоятельная органелла. Место перехода гранулярной ЭПС в агранулярную называется переходной (промежуточной, транзиторной) частью. От нее происходит отделение пузырьков с синтезированным белком и транспорт их к комплексу Гольджи. Функции гладкой ЭПС: 1. Разделение цитоплазмы клетки на отделы — компартменты, в каждом из которых идет своя группа биохимических реакций. 2. Биосинтез жиров, углеводов. 3. Образование пероксисом; 4. Биосинтез стероидных гормонов; 5. Дезинтоксикация экзо- и эндогенных ядов, гормонов, биогенных аминов, лекарств за счет деятельности специальных ферментов. 6. Депонирование ионов кальция (в мышечных волокнах и миоцитах); 7. Источник мембран для восстановления кариолеммы в телофазе митоза. ПЛАСТИНЧАТЫЙ КОМПЛЕКС ГОЛЬДЖИ. Это мембранная органелла, описанная в 1898 г. итальянским нейрогистологом К.Гольджи. Он назвал эту органеллу внутриклеточным сетчатым аппаратом из-за того, что в световом микроскопе она имеет сетчатый вид (рис. 3.4, а). Световая микроскопия не дает полного представления о строении этой органеллы. В световом микроскопе комплекс Гольджи имеет вид сложной сети, в которой ячейки могут быть связаны друг с другом или лежать независимо друг от друга (диктиосомы) в виде отдельных темных участков, палочек, зерен, вогнутых дисков. Между сетчатой и диффузной формой комплекса Гольд-жи нет принципиального различия, может наблюдаться смена форм этой оргамеллы. Еще в эпоху световой микроскопии было отмечено, что морфология комплекса Гольджи зависит от стадии секреторного цикла. Это позволило Д.Н.Насонову предположить, что комплекс Гольджи обеспечивает накопление синтезируемых веществ в клетке. По данным электронной микроскопии, комплекс Гольджи состоит из мембранных структур: плоских мембранных мешков с ампулярными расширениями на концах, а также крупных и мелких вакуолей (рис. 3.4, б, в). Совокупность этих образований называют диктиосомой. В диктиосоме находятся 5—10 мешковидных цистерн. Число диктиосом в клетке может достигать нескольких десятков. При этом каждая диктиосома связана с соседней при помощи вакуолей. В каждой диктиосоме есть проксимальная, незрелая, формирующаяся, или ЦИС-зона, -повернутая к ядру, и дистальная, ТРАНС-зона. Последняя, в отличие от выпуклой цис-поверхности, вогнутая, зрелая, обращена к цитолем- ме клетки. С цис-стороны происходит присоединение пузырьков, отделяющихся от переходной зоны ЭПС и содержащих вновь синтезированный и частично процессированный белок. При этом мембраны пузырьков встраиваются в мембрану цис-поверхности. С транс-стороны отделяются секреторные пузырьки и лизосомы. Таким образом, в комплексе Гольджи существует постоянный поток клеточных мембран и их созревание. Функции комплекса Гольджи: 1. Накопление, созревание и конденсация продуктов биосинтеза белка (происходящего в гранулярной ЭПС). 2. Синтез полисахаридов и превращение простых белков в гликопротеины. 3. Образование липонротеидов. 4. Формирование секреторных включений и выделение их из клетки (упаковка и секреция). 5. Образование первичных лизосом. 6. Формирование клеточных мембран. 7. Образование акросомы — структуры, содержащей ферменты, находящейся на переднем конце сперматозоида и необходимой для оплодотворения яйцеклетки, разрушения ее оболочек.  МИТОХОНДРИИ. Эти органеллы обеспечивают окисление органических соединений и синтез АТФ. Были открыты в 1890 году немецким ученым Р. Альтманом при помощи предложенного им метода окрашивания кислым фуксином, при этом выглядели в виде нитей и зерен (отсюда их название). Интересно, что сам Р. Альтман считал митохондрии бактериями, внедрившимися в клетку с паразитической целью, а в последующем ставшими симбионтами (симбиотическая теория происхождения митохондрий в настоящее время поддерживается многими исследователями). Размеры митохондрий составляют от 0,5 до 7 мкм, а их общее число в клетке — от 50 до 5000. Эти органеллы хорошо видны в световом микроскопе, однако информация об их строении, получаемая при этом, скудна (рис. 3.5, а). Электронный микроскоп показал, что митохондрии состоят из двух мембран — наружной и внутренней, каждая из которых имеет толщину 7 нм (рис. 3.5, б, в, 3.6, а). Между наружной и внутренней мембранами имеется щель размером до 20 нм. Внутренняя мембрана неровная, образует много складок, или крист. Эти кристы идут перпендикулярно поверхности митохондрии. На поверхности крист имеются грибовидные образования (оксисомы, АТФсомы или F,-частицы), представляющие собой АТФ-синтетазный комплекс (рис. 3.6) Внутренняя мембрана отграничивает матрикс митохондрии. Он содержит многочисленные ферменты для окисления пирувата и жирных кислот, а также ферменты цикла Кребса. Кроме того, в матриксе находятся мито-хондриальная ДНК, митохондриальные рибосомы, т-РНК и ферменты активации митохондриального генома. Внутренняя мембрана содержит белки трех типов: ферменты, катализирующие окислительные реакции; АТФ-син-тезатный комплекс, синтезирующий в матриксе АТФ; транспортные белки. Наружная мембрана содержит ферменты, превращающие липиды в реакционные соединения, участвующие затем в метаболических процессах матрикса. Межмембранное пространство содержит ферменты, необходимые для окислительного фосфорилирования. Т.к. митохондрии имеют свой геном, то они обладают автономной системой синтеза белка и могут частично строить собственные белки мембран. Функции. 1. Обеспечение клетки энергией в виде АТФ. 2. Участие в биосинтезе стероидных гормонов (некоторые звенья биосинтеза этих гормонов протекают в митохондриях). Клетки-продуценты сте роидных гормонов имеют крупные митохондрии со сложными крупными трубчатыми кристами. 3. Депонирование кальция. 4. Участие в синтезе нуклеиновых кислот. В некоторых случаях в результате мутаций митохондриалыюй ДНК возникают так называемые митохондриальные болезни, проявляющиеся широкой и тяжелой симптоматикой. ЛИЗОСОМЫ. Это мембранные органеллы, не видимые в световом микроскопе. Были открыты в 1955 году К. де Дювом при помощи электронного микроскопа (рис. 3.7). Представляют собой мембранные пузырьки, содержащие гидролитические ферменты: кислую фосфатазу, липазу, протеазы, нуклеазы и др., всего более 50 ферментов. Различают лизосомы 5 типов: 1. Первичные лизосомы, только что отделившиеся от транс-поверхности комплекса Гольджи. 2. Вторичные лизосомы, или фаголизосомы. Это лизосомы, которые соединились с фагосомой — фагоцитированной частицей, окруженной мембраной. 3. Остаточные тельца — это слоистые образования, формирующиеся в том случае, если процесс расщепления фагоцитированных частиц прошел не до конца. Примером остаточных телец могут быть липофусциновые включения, которые появляются в некоторых клетках при их старении, содержат эндогенный пигмент липофусцин. 4. Первичные лизосомы могут сливаться с погибающими и старыми органеллами, которые они разрушают. Такие лизосомы называются ауто-фагосомами. 5. Мультивезикулярные тельца. Представляют собой крупную вакуоль, в которой, в свою очередь, находятся несколько так называемых внутренних пузырьков. Внутренние пузырьки, очевидно, образуются путем отпочковывания внутрь от мембраны вакуоли. Внутренние пузырьки могут постепенно растворяться содержащимися в матриксе тельца ферментами. Функции лизосом: 1. Внутриклеточное пищеварение. 2. Участие в фагоцитозе. 3. Участие в митозе — разрушении ядерной оболочки. 4. Участие во внутриклеточной регенерации.5. Участие в аутолизс — саморазрушении клетки после ее гибели. Существует большая группа болезней, называемых лизосомными болезнями, или болезнями накопления. Они являются наследственными болезнями, проявляются дефицитом определенного лизосомального пигмента. При этом в цитоплазме клетки накапливаются непереваренные продукты 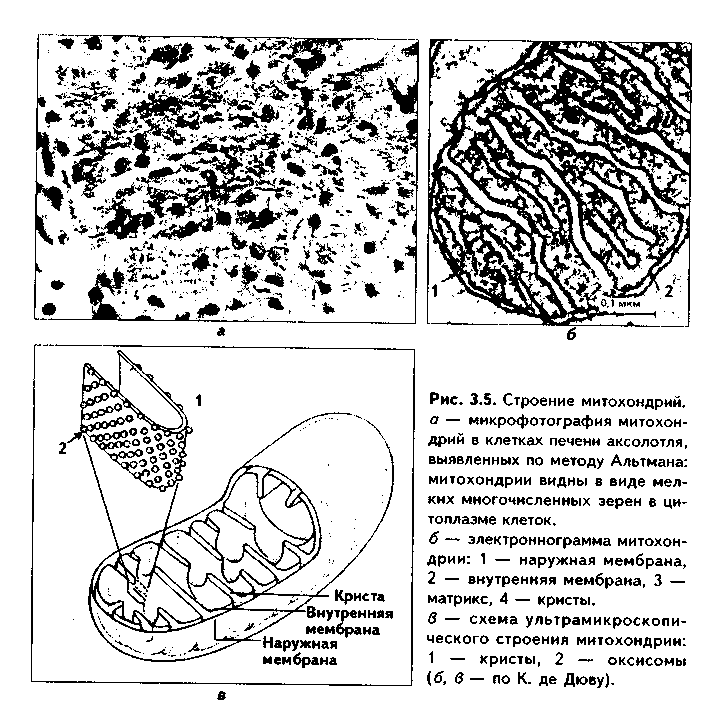 обмена веществ (гликоген, гликолиниды, белки, рис. 3.7, б,в), что ведет к постепенной гибели клетки. ПЕРОКСИСОМЫ. Пероксисомы — это оргаиеллы, напоминающие ли-зосомы, но содержащие ферменты, необходимые для синтеза и разрушения эндогенных перекисей — нероксидазу, каталазу и другие, всего до 15. В электронном микроскопе представляют сферические или эллипсоидные пузырьки с умеренно плотной сердцевиной (рис. 3.8). Образуются пероксисомы путем отделения пузырьков от гладкой ЭПС. В эти пузырьки затем миг-рируют ферменты, которые синтезируются отдельно в цитозоле или в гранулярной ЭПС 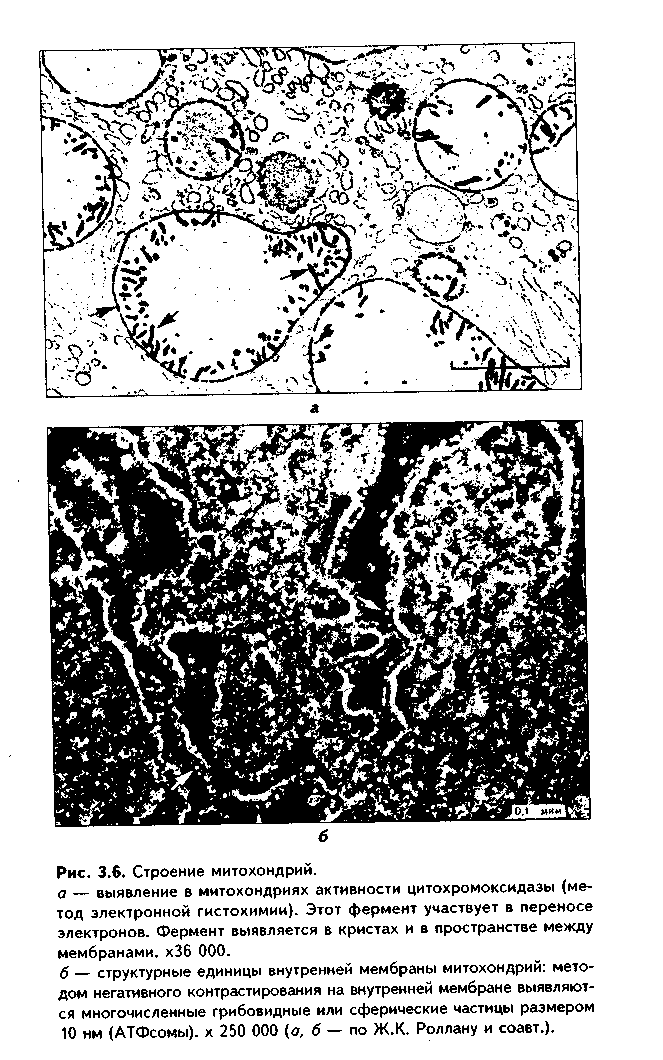 Функции пероксисом: 1. Являются, наряду с митохондриями, органеллами утилизации кислорода. В результате в них образуется сильный окислитель Н202. 2. Расщепление при помощи фермента каталазы избытка перекисей и, таким образом, защита клеток от гибели. 3. Расщепление при помощи синтезируемых в самих пероксисомах перекисей токсических продуктов, имеющих экзогенное происхождение (детоксикация). Такую функцию выполняют, например, пероксисомы печеночных клеток, клеток почек. 4. Участие в метаболизме клетки: ферменты пероксисом катализируют расщепление жирных кислот, участвуют в обмене аминокислот и других веществ.  Существуют так называемые пероксисомные болезни, связанные с дефектами ферментов пероксисом и характеризующиеся тяжелыми поражениями органов, что ведет к смерти в детском возрасте. НЕМЕМБРАННЫЕ ОРГАНЕЛЛЫ РИБОСОМЫ. Это оргаиеллы биосинтеза белка. Они состоят из двух рибонуклеоиротеидных субьединиц — большой и малой. Эти субъединицы могут соединяться вместе, при этом между ними располагается молекула информационной РНК. Есть свободные рибосомы — рибосомы, не связанные с ЭПС. Они могут быть одиночными и в виде полисом, когда на одной молекуле и-РНК находятся несколько рибосом (рис. 3.9). Вторая разновидность рибосом — связанные рибосомы, прикрепленные к ЭПС. 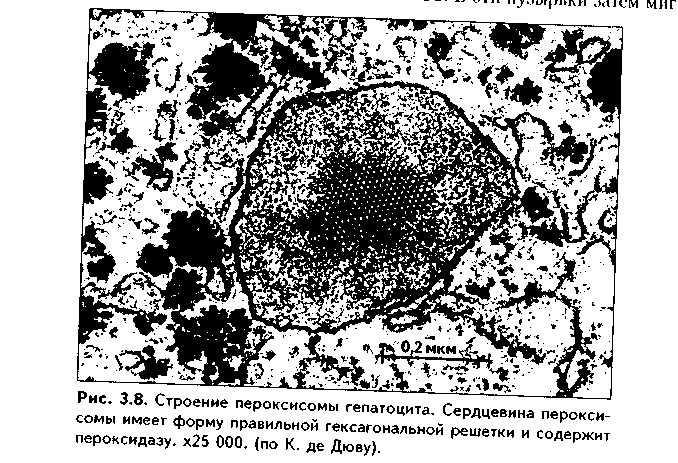 Функция рибосом. Свободные рибосомы и полисомы осуществляют биосинтез белка для собственных потребностей клетки.  Связанные на ЭПС рибосомы синтезируют белок на "экспорт", для нужд всего организма (например, в секреторных клетках, нейронах и др.). МИКРОТРУБОЧКИ. Микротрубочки являются органеллами фибриллярного типа. Они имеют диаметр 24 им и длину до нескольких мкм. Это прямые длинные полые цилиндры, построенные из 13 периферических нитей, или протофиламентов. Каждая нить образована глобулярным белком тубулином, который существует в виде двух субъединиц — аир (рис. 3.10). В каждой нити эти субъединицы располагаются поочередно. Нити в микротрубочке имеют спиральный ход. В стороны от микротрубочек отходят ассоциированные с ними молекулы белков (ассоциированные с микротрубочками протеины, или MAP). Эти белки стабилизируют микротрубочки, а также связывают их с другими элементами цитоскелета и орга-неллами. С микротрубочками связан также белок кииезин, который представляет собой фермент, расщепляющий АТФ и преобразующий энергию ее распада в механическую энергию. Одним концом кииезин связывается с определенной органеллой, а другим за счет энергии АТФ скользит вдоль микротрубочки, перемещая таким образом органеллы в цитоплазме 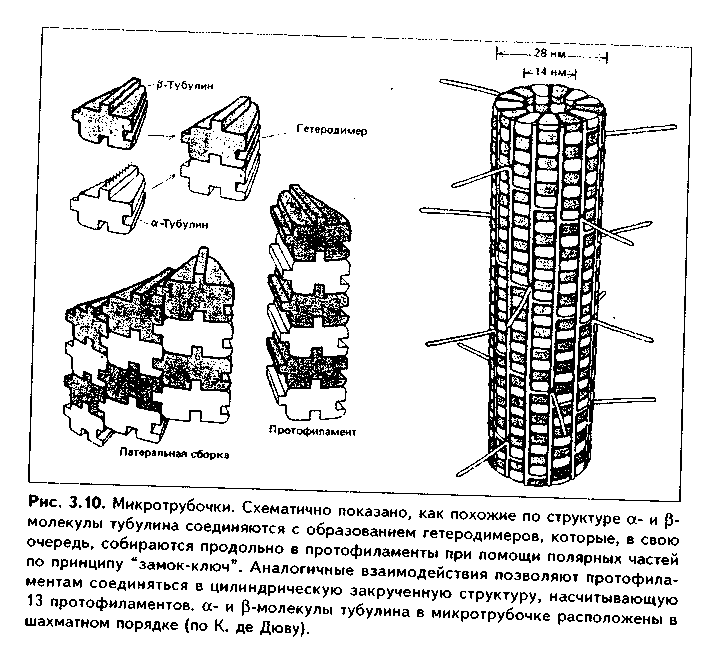 Микротрубочки являются очень динамичными структурами. Они имеют два конца: (-) и (+) — концы. Отрицательный конец является местом деполимеризации микротрубочки, тогда как на положительном конце происходит их наращивание за счет новых молекул тубулина. В некоторых случаях (базальное тельце) отрицательный конец как бы заякоривается, и распад здесь прекращается. В результате происходит увеличение размеров ресничек из-за наращивания на (+) — конце. Функции микротрубочек заключаются в следующем. 1. Выполняют роль цитоскелета; 2. Участвуют в транспорте веществ и органелл в клетке; 3. Участвуют в образовании веретена деления и обеспечивают расхождение хромосом в митозе; 4. Входят в состав центриолей, ресничек, жгутиков. Если клетки обработать колхицином, разрушающим микротрубочки цитоскелета, то клетки изменяют свою форму, сжимаются, теряют способность к делению. МИКРОФИЛАМЕНТЫ. Это второй компонент цитоскелета. Есть два вида микрофиламентов: 1) актиновые; 2) промежуточные. Кроме того, цитоскелет включает множество вспомогательных белков, которые связывают филаменты друг с другом или с другими клеточными структурами. Актиновые филаменты построены из белка актина и образуются в результате его полимеризации. Актин в клетке находится в двух формах: 1) в растворенной форме (G-актин, или глобулярный актин); 2) в полимери-зованиой форме, т.е. в виде филаментов (F-актин). В клетке существует динамическое равновесие между 2 формами актина. Как и в микротрубочках, в актиновых филаментах имеются (+) и (-) — полюсы, и в клетке идет постоянный процесс распада этих филамент на отрицательном и созидание на положительном полюсах. Этот процесс называется тредмил-лингом. Он играет важную роль в изменении агрегатного состояния цитоплазмы, обеспечивает подвижность клетки, участвует в перемещении ее органелл, в формировании и исчезновении псевдоподий, микроворсинок, протекании эндоцитоза и экзоцитоза. Микротрубочки создают каркас микроворсинок, а также участвуют в организации межклеточных включений. Промежуточные филаменты — филаменты, имеющие толщину, большую, чем у актиновых филаментов, но меньшую, чем у микротрубочек. Это самые стабильные филаменты клеток. Выполняют опорную функцию. Например, эти структуры лежат по всей длине отростков нервных клеток, в области десмосом, в цитоплазме гладких миоцитов. В клетках разного типа промежуточные филаменты отличаются по составу. В нейронах образуются ней-рофиламенты, состоящие из трех различных полипентидов. В клетках нейроглии промежуточные филаменты содержат кислый глиальный белок. В эпителиальных клетках содержатся кератиновые филаменты (тонофила-менты) (рис. 3.11).  В мышечных клетках (за исключением миоцитов сосудов) промежуточные филаменты состоят из белка десмина. В различных клетках мезенхимного происхождения, в том числе и в миоцитах сосудов, содержатся виментиновые филаменты. 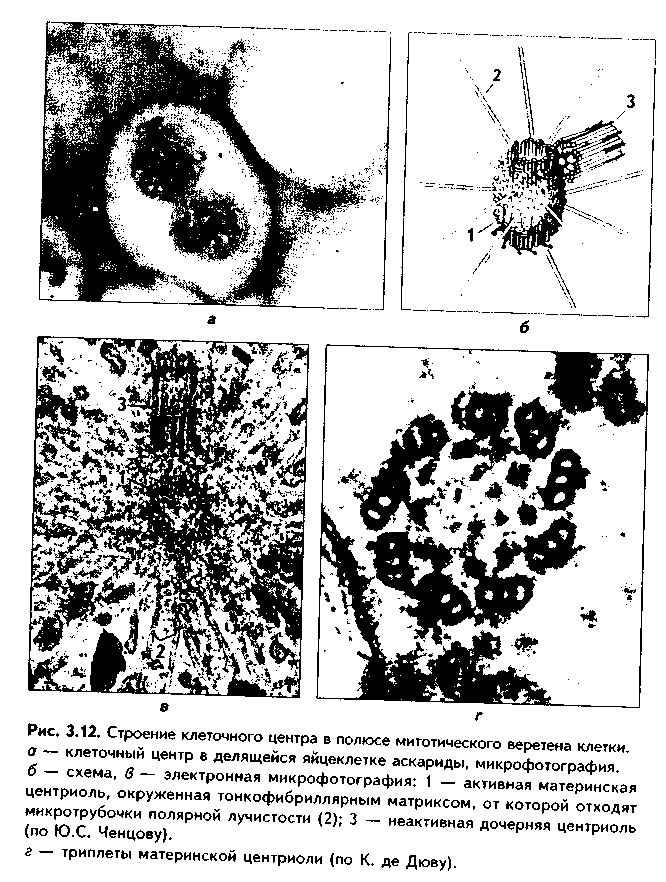 КЛЕТОЧНЫЙ ЦЕНТР (рис. 3.12). Это видимая и световой микроскоп органелла, однако ее тонкое строение позволил изучить только электронный микроскоп. В интерфазной клетке клеточный центр состоит из двух цилиндрических полостных структур длиной до 0,5 мкм и диаметром до 0,2 мкм. Эти структуры называются центриолями. Они образуют диплосому. В диплосоме дочерние центриоли лежат под прямым углом друг к другу. Каждая центриоль состоит из расположенных по окружности 9 триплетов микротрубочек, которые частично сливаются по длине. Кроме микротрубочек, в состав цептриолей входят "ручки" из белка динеина, которые соединяют соседние триплеты в виде мостиков. Центральные микротрубочки отсутствуют, и формула центриолей — (9хЗ)+0. Каждый триплет микротрубочек связан также со структурами сферической формы — сателлитами. От сателлитов расходятся в стороны микротрубочки, образуя центросферу. Центриоли являются динамичными структурами и претерпевают изменения в митотическом цикле. В неделящейся клетке парные центриоли (центросома) лежат в околоядерной зоне клетки. В S-периоде митотического цикла они дуплицируются, при этом под прямым углом к каждой зрелой центриоли образуется дочерняя центриоль. В дочерних центриолях вначале имеется только 9 единичных микротрубочек, но по мере созревания центриолей они превращаются в триплеты. Далее пары центриолей расходятся к полюсам клетки, становясь центрами организации микротрубочек веретена деления. Значение центриолей. 1. Являются центром организации микротрубочек веретена деления. 2. Образование ресничек и жгутиков. 3. Обеспечение внутриклеточного передвижения органелл. Некоторые авторы считают, что определяющими функциями клеточного центра являются вторая и третья функции, поскольку в растительных клетках центриоли отсутствуют, тем не менее и в них образуется веретено деления. РЕСНИЧКИ И ЖГУТИКИ (рис. 3.13). Это специальные органеллы движения. Они имеются в некоторых клетках — сперматозоидах, эпителиоцитах трахеи и бронхов, семявыводящих путей мужчины и др. В световом микроскопе реснички и жгутики выглядят как тонкие выросты. В электронном микроскопе установлено, что в основании ресничек и жгутиков лежат мелкие гранулы — базальные тельца, одинаковые по строению с центриолями. От базального тельца, являющегося матрицей при росте ресничек и жгутиков, отходит тонкий цилиндр из микротрубочек — осевая нить, или аксонема. Она состоит из 9 дуплетов микротрубочек, на которых находятся "ручки" из белка динеина. Аксонема покрыта цитолеммой. В центре находится пара микротрубочек, окруженная специальной оболочкой — муфтой, или внутренней капсулой. От дуплетов к центральной муфте идут радиальные спицы. Следовательно, формула ресничек и жгутиков — (9х2)+2. Основу микротрубочек жгутиков и ресничек составляет несократимый белок тубулин. Белок "ручек" — динеин — обладает АТФазной активное -гыо: расщепляет АТФ, за счет энергии которой происходит смещение дуплетов микротрубочек друг по отношению к другу. Так совершаются волнооб-ралные движения ресничек и жгутиков. Существует генетически обусловленное заболевание — синдром Карта-гснера, при котором в аксонеме отсутствуют либо динеиновые ручки, либо центральная капсула и центральные микротрубочки (синдром неподвижных ресничек). Такие больные страдают рецидивирующими бронхитами, синуситами и трахеитами. У мужчин из-за неподвижности спермиев отмечается бесплодие. МИОФИБРИЛЛЫ находятся в мышечных клетках и миосимпластах, и их строение рассматривается в теме "Мышечные ткани". Нейрофибрил-лы находятся в нейронах и состоят из нейротубул и нейрофиламентов. Их функция — опорная и транспортная. ВКЛЮЧЕНИЯ Включения — это непостоянные компоненты клетки, не имеющие строго постоянной структуры (их структура может меняться). Выявляются в клетке только в определенные периоды жизнедеятельности или жизненного цикла. 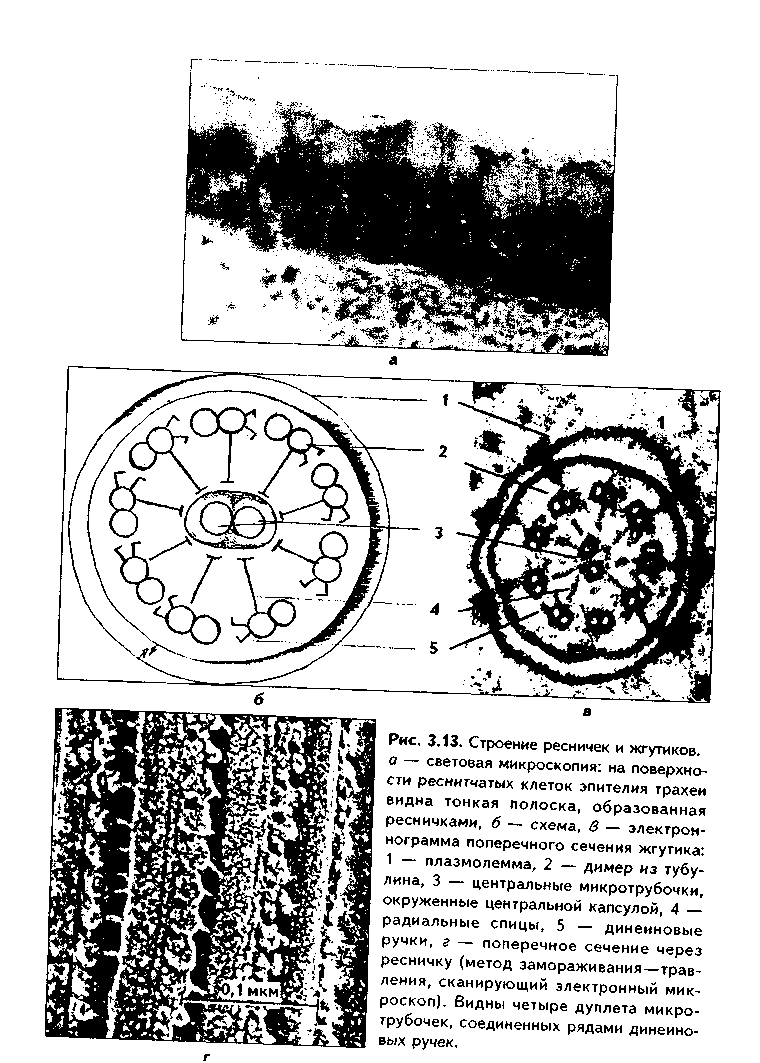 КЛАССИФИКАЦИЯ ВКЛЮЧЕНИЙ. 1. Трофические включения представляют собой депонированные питательные вещества. К таким включениям относятся, например, включения гликогена, жира. 2. Пигментные включения. Примером таких включений являются гемоглобин в эритроцитах, меланин в меланоцитах. В некоторых клетках (нервные, печеночные, кардиомиоциты) при старении в лизосомах накапливается пигмент старения коричневого цвета липофусцин, не несущий, как полагают, определенной функции и образующийся в результате изнашивания клеточных структур. Следовательно, пигментные включения представляют собой химически, структурно и функционально неоднородную группу. Гемоглобин участвует в транспорте газов, меланин выполняет защитную функцию, а липофусцин является конечным продуктом обмена. Пигментные включения, за исключением лииофусциновых, не окружены мембраной. 3. Секреторные включения выявляются в секреторных клетках и состоят из продуктов, представляющих собой биологически активные вещества и другие необходимые для осуществления функций организма вещества (включения белка, в том числе и ферментов, слизистые включения в бокаловидных клетках и др.). Эти включения имеют вид окруженных мембраной пузырьков, в которых секретируемый продукт может иметь различную электронную плотность и часто окружен светлым бесструктурным ободком. 4. Экскреторные включения — включения, подлежащие выведению из клетки, поскольку состоят из конечных продуктов обмена. Примером являются включения мочевины в клетках почки и т.д. По структуре похожи на секреторные включения. 5. Специальные включения — фагоцитированные частицы (фагосо-мы), поступающие в клетку путем эндоцитоза (см. ниже). Различные виды включений представлены на рис. 3.14. 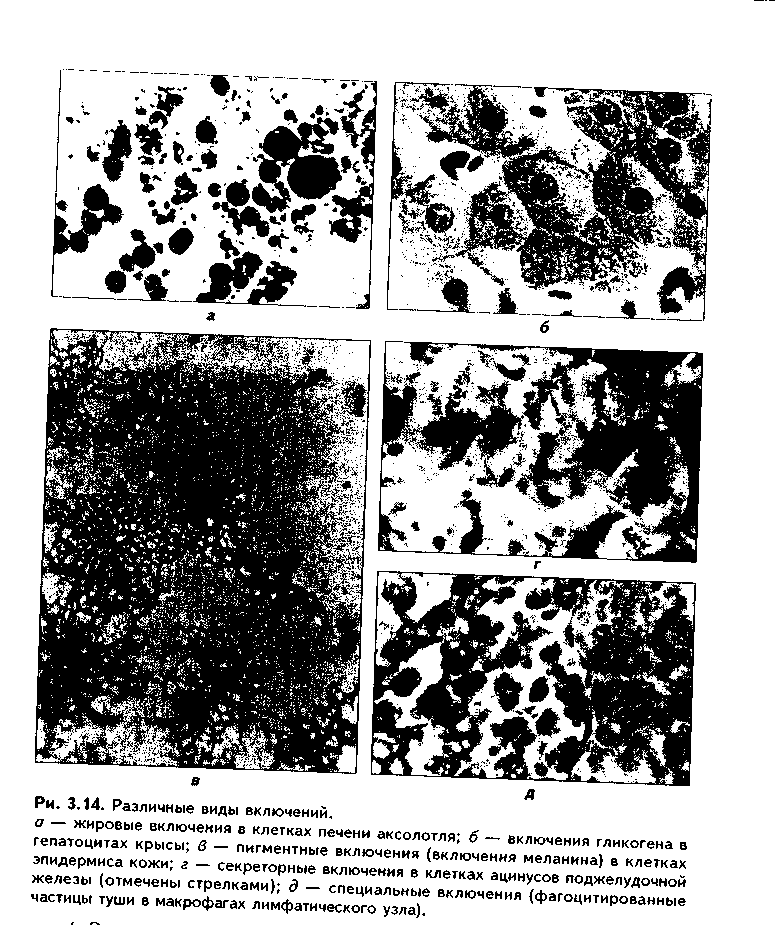 МЕЖКЛЕТОЧНЫЕ ВЗАИМОДЕЙСТВИЯ. МЕЖКЛЕТОЧНЫЕ КОНТАКТЫ Межклеточные взаимодействия — это взаимодействия клеток друг с другом. Могут быть как дистантными, на расстоянии, так и контактными. Дистантные взаимодействия осуществляются при помощи растворимых веществ, секретируемых клетками в окружающую их среду и воздействующих на другие клетки. Эти вещества называются медиаторами, или посредниками. В качестве медиаторов могут выступать гормоны, биогенные амины, антитела и многие другие биологически активные вещества, эти вещества воздействуют на репепторный аппарат клеток, с которыми взаимодействует выделившая медиатор клетка. Следовательно, дистантные межклеточные взаимодействия опосредуют действие на клетки гормонов, имеют место при иммунном ответе, эмбриональном развитии (эмбриональная индукция, см. эмбриологию) и при многих других важных клеточных реакциях. Кроме того, в многоклеточном организме все клетки связаны между собой при помощи межклеточных контактов (контактные межклеточные взаимодействия). Контактные взаимодействия состоят из нескольких фаз и включают как начальный этап дистантные взаимодействия: 1. Узнавание одной клеткой другой клетки (может быть дистантным при посредстве медиаторов и контактным при посредстве рецепторов). 2. Установление между клетками непрочных связей. 3. Формирование устойчивых межклеточных контактов. Вторая и третья фазы осуществляются при помощи молекул клеточной адгезии. Все межклеточные контакты делятся на три основных типа (рис. 3.15, 3.16): 1. Адгезионные контакты, которые механически соединяют клетки между собой. Основной тип адгезионных контактов — десмосомы. Бывают трех типов: — точечные десмосомы (пятно десмосомы). Они скрепляют клетки в отдельных местах. При этом с внутренней стороны клеточных мембран двух клеток находится электрошюплотная пластинка, связанная с сетью кератиновых микрофиламент. Эти филаменты заканчиваются в пластинке или проходят мдоль ее поверхности. Прилегающие друг к другу пластинки двух клеток соединены через межклеточное пространство волокнами из белка неизвестной природы. В межклеточном пространстве есть электронноплотный материал; — опоясывающие десмосомы (зоны десмосомы). Они идут вблизи апи-кального конца клеток по их периметру в виде полосы. Эта полоса состоит из пучков актиновых филаментов, локализующихся со стороны цитоплазмы. В межклеточном пространстве есть электронноплотный материал; — полудесмосомы. Представляют собой как бы половинку точечной десмосомы. Прикрепляют эпителиальные клетки к базальной мембране. В функционировании адгезионных контактов важную роль играют адгезионные молекулы, такие, как Е-кадгерин, дссмоколлины, десмоглеины и др. 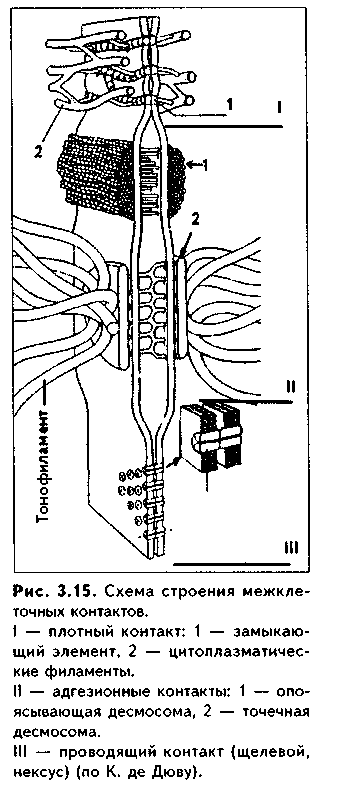  2. Плотные контакты. Это разновидность замыкающих контактов. Данный тип контактов не только механически связывает клетки друг с другом, но и препятствует прохождению между ними молекул. В плотных контактах клеточные мембраны подходят друг к другу на расстояние до 5 нм и связываются друг с другом при помощи специальных белков. 3. Проводящие контакты. В этих контактах может осуществляться передача малых молекул из одной клетки в другую. При этом мембраны двух клеток подходят друг к другу на расстояние до 3 нм и образуют каналы — коннексоны. Через коннексоны между клетками осуществляется свободный обмен низкомолекулярными веществами (электролитами, витаминами, нуклеотидами, АТФ, сахарами, аминокислотами и др.). Таким образом, этот тип контактов играет важную роль не только в механической, но и в химической коммуникации клеток. Пример таких контактов — щелевые контакты: нексусы между мышечными клетками в гладкой и сердечной мускулатуре. При этом возбуждение передается с одной клетки на другую. Второй пример — синапсы — контакты между нервными клетками. 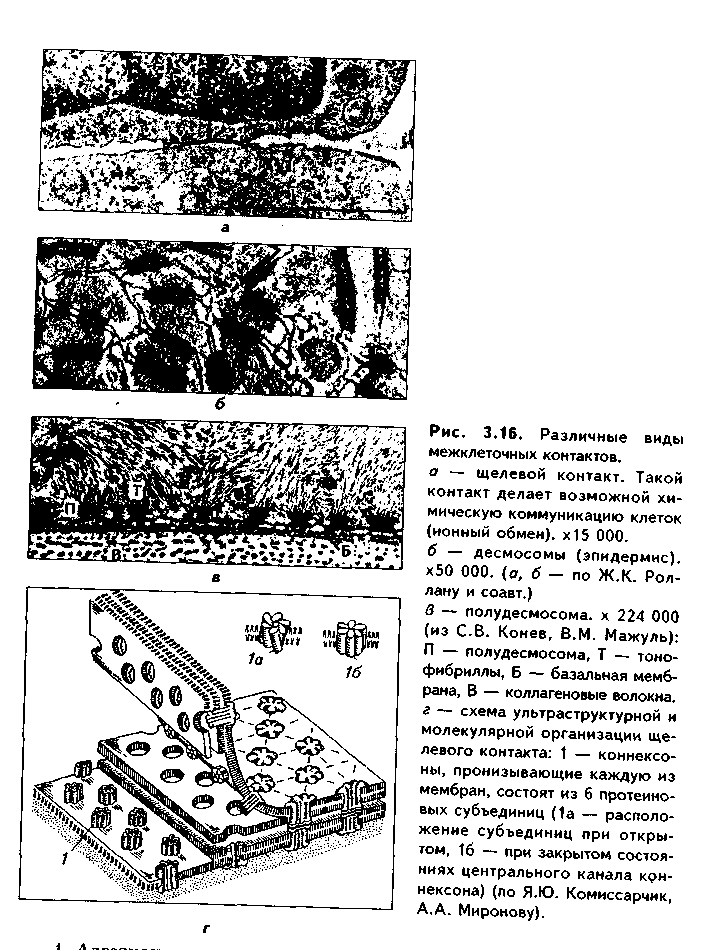 Кроме этих основных видов межклеточных контактов, выделяют также интердигитации — или межпальцевые соединения, когда цитоплазма с покрывающей ее цитолеммои одной клетки в виде пальца вклинивается в цитоплазму другой клетки и наоборот. Интердигитации резко увеличивают прочность межклеточных соединений, а кроме того, увеличивают площадь межклеточных взаимодействий, благодаря чему возрастает межклеточный обмен метаболитами. МЕХАНИЗМЫ ТРАНСПОРТА ВЕЩЕСТВ В КЛЕТКУ. ЭНДОЦИТОЗ И ЭКЗОЦИТОЗ. ОПРЕДЕЛЕНИЕ, ЗНАЧЕНИЕ, МОРФОЛОГИЧЕСКИЕ ОСНОВЫ Необходимые клетке вещества могут поступать в нее разными путями. Мелкие молекулы транспортируются путем пассивного и активного транс-порта. Пассивный транспорт не требует затраты энергии и осуществляется ио градиенту концентрации через специальные транспортные каналы (водные поры), образованные трансмембранными белками, при помощи белков-переносчиков (также трансмембранные белки). Активный транспорт осуществляется против градиента концентрации веществ и требует затраты энергии в виде АТФ. Он также осуществляется специальными белками-переносчиками. Крупные молекулы транспортируются в клетку путем эндоцитоза. Эндоцитоз — это процесс поступления в клетку макромолекул веществ из внеклеточного пространства. Он подразделяется на фагоцитоз (поступление в клетку твердых корпускулярных веществ) и пиноцитоз (поступление растворенных в воде веществ и жидкостей). В зависимости от механизмов эндоцитоза он делится на рецепторно опосредованный и рецепторно неопосредованный эндоцитоз. При рецепторно неопосредовапном эндоцитозе внеклеточный объект эндоцитоза захватывается в области инвагинации плазмолеммы клетки (рис. 3.17). Вначале фагоцитируемое вещество оказывает неспецифическое воздействие на поверхностные рецепторы клетки, которое передается на подмембранный слой микрофиламент и далее на цитоскелет. Элементы последнего вызывают впячивание цитолеммы — нишу или ямку. В нее поступает транспортируемое вещество. Ямка все более углубляется, затем края ее смыкаются, образуется иино- или фагоцитозный пузырек. Он отщепляется от основной мембраны и проникает внутрь клетки. Если пузырек содержит фагоцитируемую частицу, то он называется фагосомой, если жидкость и растворенные в ней вещества — пиноцитозным пузырьком. Фагосома может сливаться с первичными лизосомами с образованием фаголизосом. Вторая разновидность эндоцитоза опосредуется поверхностными рецензорами клетки, с которыми специфически связываются молекулы объекта эндоцитоза (лиганда). При этом происходит более быстрое поглощение ли-гапда в комплексе с рецепторами клетки. Очень часто при рецеиторпооносредованном эндоцитозе рецепторы клетки осуществляют кэппинг, т.е. мигрируют латерально и накапливаются в области образующихся эпдоцитозных ямок. Одновременно вокруг эн-доцитозных ямок накапливается белок клатрин, образуя сстевидную оболочку. Так формируются окаймленные пузырьки. Содержимое этих пузырьков может подвергаться превращению внутри клетки только после утраты клатриновой оболочки. Без этого пузырек не может сливаться с лизосомами, другими клатриновыми пузырьками, т.е. как бы депонируется в клетке. Окаймленные пузырьки используются для транспорта иммуноглобулинов, желточных включений в овоците, факторов роста, липопротеи-нов низкой плотности и ряда других веществ. Они являются своего рода аккумуляторами клеточных рецепторов, т.к. в них происходит предпочтительное концентрирование реценторных белков. Циторецепторы, аккумулированные в окаймленных пузырьках, служат своего рода депо рецепторов, поскольку их мембраны могут при необходимости встраиваться в ци-толемму. Благодаря этому окаймленные пузырьки позволяют одновременно подвергнуть эндоцитозу большое количество молекул лиганда при экономичном расходовании цитомембран. Примером рецепторно опосредованного эндоцитоза может быть фагоцитоз лейкоцитом окруженных антителами бактерий (рис. 3.18). В данном случае иммуноглобулины (опсонины) используются как рецепторы лиганда, с которыми комплементарно взаимодействуют поверхностные рецепторы фагоцита. Экзоцитоз — явление, в определенной степени противоположное эндоцитозу, "эндоцитоз наоборот" (рис. 3.17, 3.18). 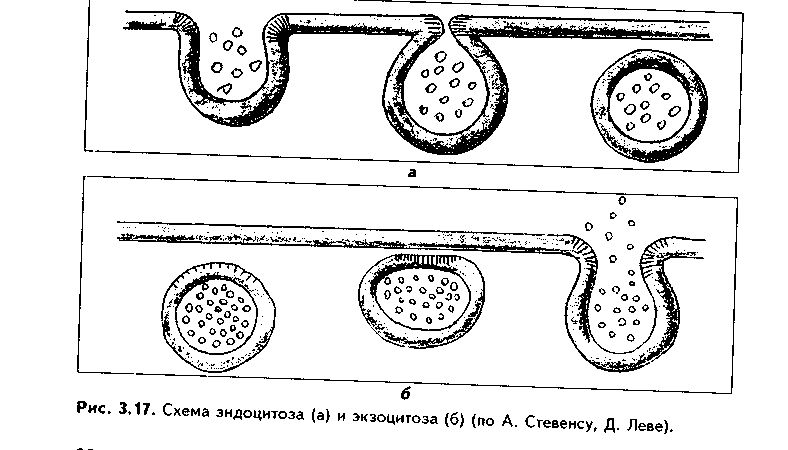 Это выделение клеткой продуктов секреции или конечного обмена. В случае секрета секреторные гранулы, окруженные мембраной, полученной в комплексе Гольджи, передвигаются в результате сокращения цитоскелета к цитолемме, сливаются с ней. Затем секреторный пузырек раскрывается, и секрет оказывается за пределами клетки. Экзоцитоз лежит в основе так называемой мерокрино-вой секреции желез (см. ЖЕЛЕЗЫ). Выделяемые из клетки путем экзоцитоза вещества могут оставаться на ее оболочке в виде рецепторов, могут входить в состав межклеточного вещества либо после попадания в межклеточную жидкость играть роль сигнальных молекул (гормоны и др.). Пиноцитозные пузырьки могут оставаться в клетке, но могут мигрировать на противоположную сторону клетки и там отрываются с выделением их содержимого. Это явление называется трансцитозом и служит для транспорта веществ. Следовательно, трапсцитоз совмещает к себе эндоцитоз и экзоцитоз. Особенно интенсивно он протекает в клетках кровеносных и лимфатических сосудов — эндотелиоцитах. ЦИТОСКЕЛЕТ. Цитоскелет — совокупность опорно-сократительных структур клетки, система идущих в разных направлениях и образующих трехмерную сен. актиновых филамент, микротрубочек, микротрабекул и промежуточных филамент (см. рис. 3.10, 3.11). Актиновые филаменты лабильны, могут быстро распадаться и снова собираться (тредмиллинг). В результате изменяется форма клеток и обеспечивается их подвижность. Микротрубочки также обладают способностью распадаться и снова собираться. Они имеете с промежуточными филаментами выполняют опорную функцию. Промежуточные филаменты по диаметру тоньше микротрубочек, но толще актиновых микрофиламент. Микротрабекулы как элемент иитоскелета видны только при высоковольтной электронной микроскопии и являются наименее изученным компонентом иитоскелета. Имеют толщину 2—10 нм. Микротрабекулы формируют в клетке нежную сеть, которая интегрирует все другие элементы иитоскелета и плазмолемму. В узлах микротрабекулярной сети находятся рибосомы и полисомы. Химический состав микротрабекул не выяснен. Цитоскелет, с одной стороны, связан с подмембранным слоем клеточной оболочки и интегральными белками плазмолеммы, с другой — с многими органеллами в цитоплазме и с ядром. Внеклеточные сигналы, в том числе и от молекул межклеточного матрикса (фибронектин), действуя через циторецепторы, могут реализоваться цитоскелетом через подмембран-ный слой. Функции иитоскелета: 1. Опорная. Создает жесткий каркас клетки. 2. Регуляция вязкости и формы клетки, обеспечение ее движения. 3. Участие в эндо- и экзоцитозе и связанных с ними клеточных процессах (пиноцитоз, фагоцитоз, секреция и др.). 4. Участие в цитотомии при митозе. При этом задействованы актиновые филаменты, которые концентрируются в области цитотомии, сокращаются и образуют перетяжку, углубляющуюся до полного отделения клеток друг от друга. 5. Внутриклеточный транспорт макромолекул и органелл. 6. Обеспечение латеральной подвижности рецепторных белков в ли-пидном бислое цитолеммы и кэппинга, имеющего значение в ответной реакции клетки на разражители. 7. Промежуточные филаменты являются показателем тканевой принадлежности клеток, поскольку клетки каждого тканевого типа имеют свой специфический белковый состав. Эпителиальные клетки содержат кератиновые филаменты, мышечные — в основном десминовые, соединительнотканные клетки — виментиновые, нервные — иейрофиламенты, глиаль-ные клетки — глиальные филаменты, содержащие кислый фибриллярный глиальный белок. 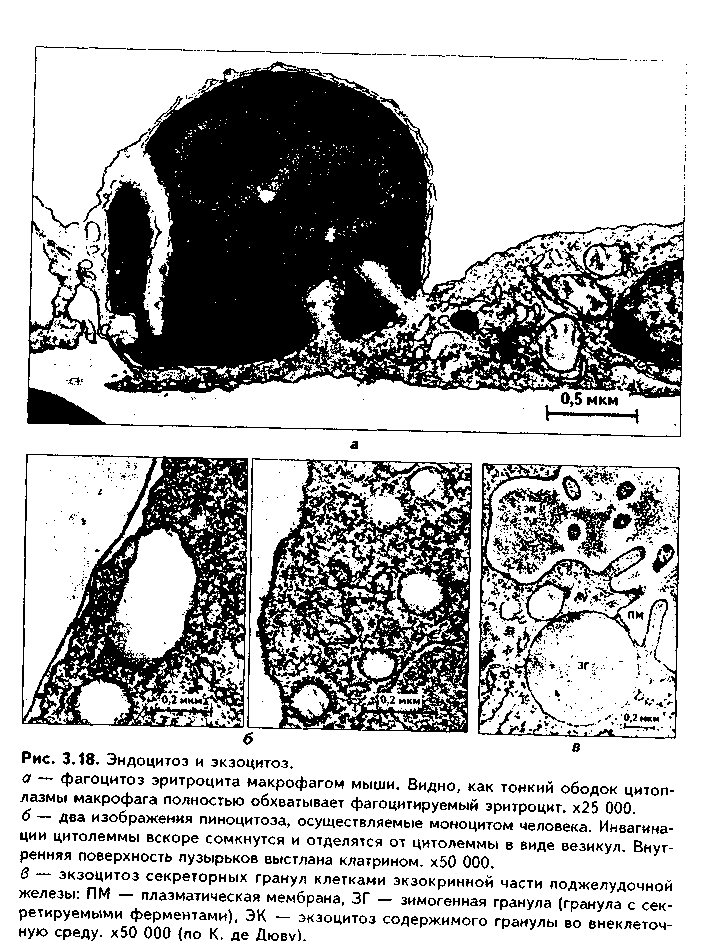 ВНЕКЛЕТОЧНЫЙ (ЭКСТРАЦЕЛЛЮЛЯРНЫЙ) МАТРИКС (ВМ). Внеклеточный матрикс — это вещество, находящееся между клетками. В соединительных тканях межклеточный матрикс является одним из тканевых элементов и называется межклеточным веществом, которое состоит из волокон (коллагеновые, эластические, ретикулярные) и основного, или аморфного вещества (см. СОЕДИНИТЕЛЬНЫЕ ТКАНИ). Аморфное вещество состоит из воды и различных макромолекул: белков, углеводов (гликозаминогликаны и другие), комплекса белков с гликозаминог-ликанами (гликопротеины, протеогликаны), а также других веществ. В эпителиальной ткани внеклеточный матрикс слабо выражен, состоит в основном из аморфного вещества. Особой формой межклеточного матрикса в эпите-лиях являются базальные мембраны. Их строение будет рассмотрено в теме "Эпителиальная ткань". Одними из наиболее важных молекул внеклеточного матрикса, играющих роль в межклеточных взаимодействиях и во взаимодействиях "клетка — внеклеточный матрикс", являются ламинин, фибронектин и нидоген/эн-тактин. Они взаимодействуют с рецепторами на поверхности клеток — ("интегринами"), которые через внутриклеточные белки таллин, винкулин и а-актинин передают информацию на актиновые филаменты цитоскелета. Поэтому механические, физические и химические изменения в ВМ ведут к изменению функций клеток. Существует и обратный путь передачи информации — от внутриклеточных структур на ВМ. Функции внеклеточного матрикса: 1. Опорная. 2. Обеспечение обменных процессов и поступление в клетку веществ. 3. Регуляторная. Осуществляет регуляцию деятельности клеток. 4. Морфогенетическая, т.е. ВМ принимает участие в формировании тканевой архитектоники. Кроме того, ВМ участвует в гисто- и органогенезе, канцерогенезе и метастазировании опухолевых клеток, заживлении ран. 5. Транспортная. ВМ обеспечивает поступление к клетке необходимых регуляторных и питательных веществ. ВЗАИМОДЕЙСТВИЕ СТРУКТУР КЛЕТКИ В ПРОЦЕССЕ ЕЕ МЕТАБОЛИЗМА НА ПРИМЕРЕ СИНТЕЗА БЕЛКОВЫХ И НЕБЕЛКОВЫХ ВЕЩЕСТВ Все клеточные органеллы и структуры тесно связаны между собой при выполнении клеткой ее функций. Это можно продемонстрировать на примере синтеза клеткой белковых и небелковых секретов. При синтезе белковых веществ наблюдается следующая цепь событий: 1. Происходит транскрипция ДНК и образуется и-РНК. 2. В ядрышке образуются рибосомы, которые поступают в цитоплазму. 3. В случае синтеза белка на экспорт рибосомы присоединяются к ЭПС. 4. В митохондриях образуется АТФ, необходимая для биосинтеза белка. 5. На гранулярной ЭПС синтезируется и частично процессируется полипептидная цепь. 6. Она поступает в комплекс Гольджи, где превращается в сложный белок, а также упаковывается в мембранный пузырек. Образуются секреторные гранулы. 7. Секреторные гранулы в результате сокращения цитоскелета движутся к поверхности клетки и выделяются путем экзоцитоза. При синтезе небелковых веществ происходят следующие события: 1. Происходит транскрипция ДНК с образованием и-РНК. В ядрышке образуется рибосомальная РНК и осуществляется сборка предшественников рибосом, которые поступают в цитоплазму. 2. На свободных рибосомах в цитоплазме синтезируются ферменты биосинтеза небелковых веществ. 3. Они переходят в гиалоплазму или в гладкую ЭПС, где синтезируются небелковые вещества — углеводы, липиды. 4. Эти вещества поступают в комплекс Гольджи, там окружаются мембранами, а далее формируются секреторные гранулы, выделяемые из клетки. Таким образом, все компоненты клетки тесно функционально связаны между собой. Следует отметить также, что в клетке существует постоянный поток клеточных мембран — рециклинг (оборот мембран, мембранный конвейер). Белковые компоненты мембран синтезируются на рибосомах, липид-ные и углеводные — в цитозоле и в гладкой ЭПС. После сборки они включаются в ЭПС, от которой могут отделяться в виде пузырьков и присоединяться к комплексу Гольджи, входя уже в состав его мембран. Транс-сторона комплекса Гольджи отделяет секреторные пузырьки, которые затем выделяют свое содержимое путем экзоцитоза. При этом их мембрана встраивается в цитолемму ("плюс-мембрана"). С другой стороны, при эндоцитозе часть цитолеммы идет на построение оболочки эндо-сом ("минус-мембрана"). Оба процесса мембранного конвейера клетки строго уравновешены, и обычно не происходит ни уменьшения, ни увеличения площади поверхности клетки. |