МЯДЕЛЕЦ ОБЩ.ГИСТ.. Литература для медицинских вузов все факультеты о. Д. Мяделец основы цитологии, эмбриологии и общей гистологии
 Скачать 4.75 Mb. Скачать 4.75 Mb.
|

|
Глава 6 ГИСТО- И ОРГАНОГЕНЕЗ. РАЗВИТИЕ ОСНОВНЫХ ОРГАНОВ И СИСТЕМ ПЛОДА НА 4—8-й НЕДЕЛЯХ ЭМБРИОГЕНЕЗА. ПЛАЦЕНТА. ПОНЯТИЕ О ФУНКЦИОНАЛЬНОЙ СИСТЕМЕ «МАТЬ—ПЛОД» ГИСТОГЕНЕЗ И ОРГАНОГЕНЕЗ. РАЗВИТИЕ ОСНОВНЫХ ОРГАННЫХ СИСТЕМ НА 4—8-й НЕДЕЛЯХ ЭМБРИОГЕНЕЗА Источником развития тканей являются эмбриональные зачатки. В свою очередь, эмбриональные зачатки развиваются из зародышевых листков и процессе их дифференцировки. В результате формируется осевой комплекс зачатков (см. гл. 5). Процесс образования тканей в эмбриогенезе из тканевых зачатков называется эмбриональным гистогенезом. Механизмы гистогенеза достаточно сложны и включают следующие компоненты: 1. Деление клеток. В результате деления клеток зачатка происходи! нарастание клеточного материала, объема зачатка, достижение им критической массы, что запускает дальнейшие гистогенетические процессы. Основным видом деления клеток в ходе гистогенеза является митоз. Он может быть стволовым, асимметричным и дифференцирующим, или квантальным. При стволовом митозе из одной материнской стволовой клетки образуются две дочерние стволовые клетки. Для асимметричного митоза характерно то, что из двух дочерних клеток одна является стволовой, а вторая вступает на путь дифференцировки. При квантальном митозе обе дочерние клетки отличаются от стволовых, поскольку уже приступили к дифференцировке. 2. Рост клеток. Наряду с митозом рост клеток приводит к увеличению общей массы зачатка ткани. В его основе лежат гипертрофия и гиперплазия клеточных органелл, накопление включений. 3. Запрограммированная гибель клеток, или апоптоз. По своему значению клеточная гибель не менее важна для гистогенетических процессов, чем деление клеток. В результате аиоптоза регулируется число клеток в разнимающейся ткани, происходит ее перестройка, исчезают рудиментарные за-читки, элиминируются мутировавшие и дефектные клетки. Любопытно, что 1 некоторых случаях в ходе гистогенеза сразу образуется заведомо больше Меток, чем их необходимо для развития ткани, и это создает определенный материальный базис гистогенеза. В последующем лишние клетки погибают, причем уничтожаются менее полноценные или дефектные клетки. Особенно •то явление выражено в нервной ткани, где в ходе гистогенеза гибнет от 50 до 85 % всех нейронов. 4. Миграция клеток. Различают пассивную и активную миграцию клеток. Пассивная миграция — миграция в результате давления соседних клеток. Активная миграция клеток происходит за счет работы внутриклеточных сократительных структур, связанных через подмембранный слой с поверхностными рецепторами. 5. Адгезия клеток и межклеточные взаимодействия (механизмы клеточной адгезии — см. цитологию). Для образования ткани необходимо, чтобы клетки зачатка совершили миграционные процессы, а затем сформировали клеточные ансамбли. Инициация миграции связана с потерей клетками зачатка адгезионных молекул (эта ситуация определяется как конец адгезии — начало миграции). После начала миграции клеточная адгезия контролирует миграцию клеток: мигрирующие в ходе гистогенеза клетки узнают на поверхности других клеток или во внеклеточном мат-риксе адгезионные молекулы, что обеспечивает целенаправленность миграции. После завершения миграции начинается процесс формирования нужных клеточных ансамблей. При этом в завершивших миграцию клетках вновь появляются молекулы адгезии, и между клетками устанавливаются взаимодействия (конец миграции — начало адгезии). 6. Детерминация (процесс определения пути, программы развития эмбриональных зачатков в направлении той или иной дефинитивной ткани). Механизм детерминации связан со стойкой репрессией одних и дерепрес-сией других генов, необходимых для развития клеток будущей ткани в нужном направлении. 7. Дифференцировка — стойкое структурно-функциональное изменение ранее однородных клеток, приобретение ими специфических черт строения для выполнения специфических функций. Молекулярно-генетические основы дифференцировки — транскрипция, сплайсинг РНК, ее процессинг, трансляция, т.е. синтез специфических и-РНК и на них — специфических белков. Морфологической основой дифференцировки является образование из специфических белков специфических клеточных органелл. 8. Эмбриональная индукция. Эмбриональная индукция — это направление гистогенетических процессов в нужное русло путем выделения одним зачатком веществ — индукторов, действующих на другой зачаток. В качестве эмбриональных индукторов могут выступать не только химичесьь индукторы, биологически активные вещества и гормоны (вторичные ни дукторы), но и самые обычные факторы: питательные вещества, уровен:. рН, концентрация электролитов, кислорода и др. (первичные индукторы) Органогенез — процесс образования органов и систем органов из умГ> риональных зачатков. Этот процесс протекает обычно параллельно с пк тогенезом, т.е. с образованием тканей в составе будущих органов, и от/и лить два процесса друг от друга невозможно. В процессе органогенез.! организм зародыша разделяется на относительно независимо развивают и сея местные системы, дающие орган. Многие механизмы гистогенеза и ор ганогенеза являются общими. ОБЩИЕ МЕХАНИЗМЫ ОРГАНОГЕНЕЗА А. Клеточные процессы. 1. Клеточное размножение — непременная предпосылка органогенеза. 2. Формообразующую роль играет клеточная гибель. Например, паль цевые фаланги разъединяются потому, что клетки в промежутках межд\ ними гибнут. Гибель клеток лежит в основе кавитации — образования по лостей в полых органах и канальцах. 3. Формообразующая роль клеточных движений. В органогенетически.ч процессах участвуют всевозможные типы клеточных движений. За счет перемещения клеток поставляется клеточный материал в те или иные отде лы зародыша, что приводит к утолщениям и изгибам клеточных пластов необходимым в процессе формирования органов. 4. Избирательная сортировка клеток (сегрегация). В ходе морфогенез;: клетки движутся не с абсолютной точностью, возможны ошибки, и тогда "наводится" порядок путем сортировки. Явления клеточной сортировки определяются контактными взаимодействиями клеток через посредство рецепторов. Эти же механизмы поддерживают структуру тканей в дефинитивных органах, а их утеря приводит к злокачественному росту. Б. Морфогенетические преобразования в зачатках. Клеточные процессы приводят к морфологическим преобразованиям в зачатках: 1. Изгиб клеточного пласта. Имеет место при развитии органов из эпителиальных пластов. Все формообразование в ЦНС, органах пищеварения и ряде других органов в первую очередь сводится к последовательным изгибам. 2. Утолщение некоторого участка эпителиального пласта. По таком\ механизму (из утолщения эпителия — плакод) развиваются органы обоняния, слуха, хрусталик, некоторые органы полости рта. В мезенхимных зак- ладках наблюдается аналогичный процесс сгущения клеток, предшествующий, например, закладке хряща или кости. 4. Противоположный предыдущему процесс — разрежение клеток, или кавитация, что лежит, как упоминалось, в основе образования полых органов. В. Межзачатковые индукционные взаимодействия. В развитии органов важную роль играют межзачатковые индукционные изаимодействия. Например, зачаток глазного бокала, влияя на эктодерму, стимулирует образование из нее хрусталика как части будущего органа зрения. На более поздних стадиях развития глазной бокал индуцирует преобразование кожи в роговицу. Осуществляются межзачатковые индукционные взаимодействия при помощи первичных и вторичных индукторов. Г. Межтканевые взаимодействия. Осуществляются на основе индукционных взаимодействий при помощи третичных индукторов, воздействующих на уже детерминированный клеточный материал. Наиболее частыми являются взаимодействия энтодерма—мезенхима и эктодерма—мезенхима. Именно эти пары зародышевых листков дают ткани, которые, тесно взаимодействуя друг с другом, дают большинство органов. Д. Нервные, эндокринные и иммунные влияния. На определенном этапе органогенеза образование органов становится невозможным без участия регуляторных механизмов. Таковыми являются нервные, эндокринные и иммунные регуляторные влияния. РАЗВИТИЕ ОСНОВНЫХ ОРГАННЫХ СИСТЕМ НА 4—8-й НЕДЕЛЯХ ЭМБРИОГЕНЕЗА Образование основных закладок органов происходит с 4-й по 8-ю неделю эмбриогенеза. В это время из нервной трубки формируются головной и спинной мозг. При развитии головного мозга передняя часть нервной трубки превращается вначале в три мозговых пузыря, а затем формируются пять мозговых пузырей, из которых образуются пять отделов головного мозга. Мозговые пузыри просвечиваются через кожу. На 6-й неделе эмбриогенеза наибольшего развития достигают передний и промежуточный мозг. Происходит закладка глаз и дифференциропка нейронного слоя сетчатки. Зачатки глаз видны в виде небольших круглых пятнышек. Позади них располагаются зачатки трех пар жаберных карманов. Просвечивается закладка щитовидной железы. На 5-й неделе эмбриогенеза происходит дальнейшая дифференциров-ка и усложнение строения органов сердечно-сосудистой системы. Закладка сердца и сосудов образуется на 3-й неделе эмбриогенеза, но формирование основных оболочек сердца и сосудов, усложнение их строения, дифферен цировка кардиомиоцитов и формирование проводящей системы сердцг происходит в течение второго месяца эмбриогенеза. В это же время идет формирование основных эндокринных органов. На 4—6-й неделях закладываются все органы желудочно-кишечного тракта, а печень и поджелудочная железа, которые начали образовываться в конце третьей недели эмбриогенеза, подвергаются дальнейшему развитию. На втором месяце эмбриогенеза закладывается вторичная почка, а затем происходит ее дальнейшее развитие. К восьмой неделе завершается дифференцировка гонад. Органы иммунной системы и кроветворения начинают закладываться несколько позднее, чем другие органы. Так, лимфоузлы впервые появляются только к концу восьмой недели, селезенка формируется к концу пятой недели. Тимус закладывается в конце первого месяца эмбриогенеза, но лимфоцитами заселяется только к концу второго месяца. Для того, чтобы оценить, какие изменения происходят с зародышем с 4-й по 8-ю неделю эмбриогенеза, рассмотрим основные черты анатомического строения его в эти сроки (рис 6.1). В конце 4-й — начале 5-й недели эмбриогенеза зародыш человека с трудом отличим от зародышей других высших млекопитающих, находящихся на аналогичных стадиях развития. Его тело имеет длину 3,5 мм, зародыш находится на стадии 35 пар сомитов. Тело зародыша изогнуто в вентральном направлении, особенно в области головы и хвоста. На границе между головой и телом постепенно обозначается резкий шейный изгиб. Благодаря этому головной конец постепенно приближается к резко выраженному сердечному выступу и упирается в него. Хорошо видны зачатки рук в виде плавников, а зачатки ног только начинают развиваться. Головной конец значительно больше хвостового, шея не выражена. Имеется хвост. К концу 8-й недели эмбриогенеза зародыш приобретает несомненные человеческие черты. Полностью формируются черты лица, редуцируется хвост. Конечности удлиняются, сформированы все их отделы. 8-недельный зародыш имеет длину около 40 мм и весит около 5 г. К этому времени происходит уплощение висцеральных дуг и обособляется шея. Голова становится круглой. Образуются наружное ухо и наружные части носа. Глаза смещаются кпереди и сближаются. Хорошо развиты пальцы. В переднем отделе мозга начинается усиленный рост больших полушарий. Сформированы все внутренние органы. 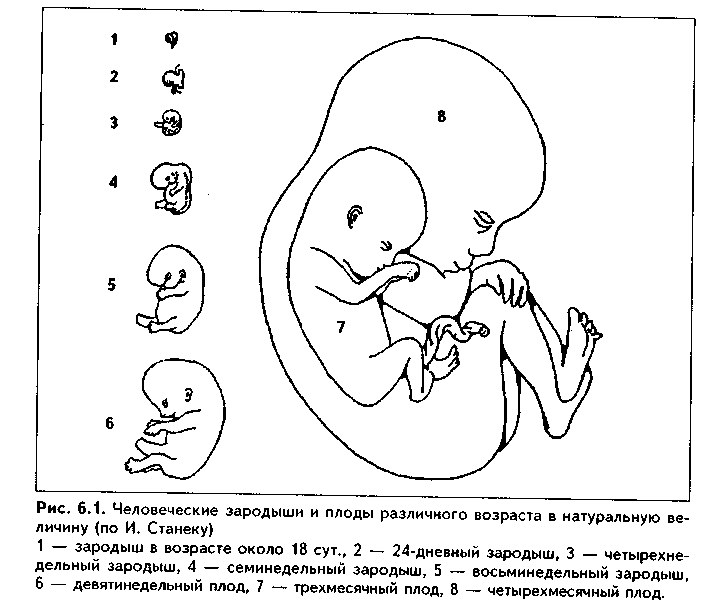 Таким образом, к концу восьмой недели эмбриогенеза завершается формирование основных органных систем зародыша. К этому моменту завершается зародышевый период эмбриогенеза и начинается ПЛОДНЫЙ ПЕРИОД, который длится до конца беременности. ПРОВИЗОРНЫЕ ОРГАНЫ. ОБРАЗОВАНИЕ, СТРОЕНИЕ, ФУНКЦИИ Провизорные органы — это временные органы зародыша и плода, обеспечивающие его нормальное развитие. Источником их развития являются внезародышевые части зародышевых листков. Некоторые провизорные органы (аллантоис, желточный мешок) существуют непродолжительное время и после выполнения своих функций подвергаются редукции. Другие (хорион и образующаяся из него плацента, амнион, пупочный канатик) существуют до момента рождения.Первыми из провизорных органов образуются амнион и желточный мешок. Как уже отмечалось (см. гл. 5 и рис. 5.10), вначале из эпибласта выселяются клетки, которые заполняют всю полость бластоцисты и образуют первичную мезенхиму. В первичной мезенхиме образуются две полости: над эпибластом и под гипобластом. За счет размножения клеток эпибласта и гипобласта эти полости обрастают клетками первичной эктодермы и первичной энтодермы. В результате формируются два п> зырька: амниотический и желточный. Их стенки образуют амнион и желточный мешок. АМНИОН. Амнион образуется из первичной внезародышевой эктодер мы и первичной мезенхимы, из которых на ранних этапах эмбриогенеза и состоит его стенка. Внезародышевая эктодерма превращается в амнионi ческий эпителий — однослойный плоский, затем цилиндрический, места ми многорядный эпителий. Эпитслиоциты лежат на базалыгой мембране и на апикальной поверхности имеют микроворсинки. Внезародышевая мезодерма превращается в соединительнотканную основу амниона, которая делится на два слоя: компактный, образованный плотной соединительной тканью, находится сразу под эпителием, и губчатый, представленный ст\ денистой соединительной тканью, лежащий глубже и обращенный к хори альной пластинке.  ФУНКЦИИ. 1. Амнион выделяет жидкость и формирует водную оболочку вокруг зародыша, которая защищает его от неблагоприятных воздействий внешней среды, в первую очередь от механических — защит но-механическая и секреторная функции. 2. Всасывательная функция -обратное всасывание околоплодных вод. Постоянные уравновешенные процессы секреции и всасывания обеспечивают обмен околоплодных вод. 3. Регуляторная функция. Плод постоянно заглатывает определенное ко личество секретируемых амнионом околоплодных вод, которые стимулируют эмбриогенез и деятельность желудочно-кишечного тракта плода. 4. Выделительная функция. В околоплодные воды плод выделяет мочу и с ней конечные продукты обмена. Эти же продукты выделяются с поверхности кожи. 5. Эндокринная функция. На поздних этапах эмбриогенеза амнион вырабатывает простагландины, стимулирующие родовую деятельность. Прием беременной женщиной накануне родов салицилатов, угнетающих синтез простагландинов, способствует перенашиванию беременности. ЖЕЛТОЧНЫЙ МЕШОК. Желточный мешок полностью формируется на 11-е сутки эмбриогенеза. Источником его развития являются внезародышевая энтодерма и внезародышевая эктодерма (рис. 5.10, 6.2). Иногда желточный мешок рассматривают как вынесенную за пределы зародыша часть первичной кишки. После образования туловищной складки желточный мешок отделяется от кишечной трубки, но остается связанным с ней желточным стебельком. В дальнейшем желточный мешок смещается в пространство между хорионом и амнионом и включается в состав пупочного канатика, где сохраняется в виде узкой трубки. Функционирует до 7—8-й недель эмбриогенеза, а затем подвергается обратному развитию. ФУНКЦИИ. 1. Трофическая. Желточный мешок не содержит, как это наблюдается у птиц, больших запасов питательных веществ, однако опре- деленное их количество в нем имеется. Поэтому на ранних этапах эмбриогенеза желточный мешок обеспечивает питание зародыша. Его клетки содер жат ферменты, расщепляющие желточные массы. Кроме того, желточный мешок на ранних этапах эмбриогенеза способен активно всасывать питательные вещества из расположенных близко к нему сосудов матки до развития хориона. 2. Дыхательная. Кроме питательных веществ желточный мешок поглощает из сосудов матки кислород. 3. Продукция первичных половых клеток. В энтодерме желточного мешка на 3-й неделе эмбриогенеза образуются первичные половые клетки, которые в последующем мигрируют в закладки гонад. 4. Кроветворная. На 3-й неделе эмбрионального развития в мезенхиме желточного мешка образуются первичные клетки крови и первичные кровеносные сосуды. Кроветворную функцию желточный мешок выполняет и промежуток времени с 3-й по 7—8-ю недели эмбриогенеза (внезародышевый период эмбрионального гемопоэза). АЛЛАНТОИС. Образуется как колбасовидное выпячивание вентральной стенки энтодермы задней кишки в амниотическую ножку на 16-е сутки эмбриогенеза, т.е. после гаструляции (рис. 6.2). Снаружи он покрывается внезародышевой мезенхимой амниотической ножки. Таким образом, аллан-тоис состоит из двух слоев: внезародышевой энтодермы и внезародышевой мезенхимы. Дистальная часть аллантоиса быстро растет и превращается в соединенный с кишкой при помощи ножки мешок. У человека аллаитоис но достигает крупных размеров и существует до 2-го месяца эмбриогенеза. При формировании пупочного канатика аллантоис включается в его состав, где затем подвергается редукции. ФУНКЦИИ. 1. Участие в формировании сосудистой сети плаценты. Является проводником кровеносных сосудов из желточного мешка во вторичные ворсины. 2. Гистогенетическая — проксимальная часть аллантоиса идет на образование части переходного эпителия мочевого пузыря, и нарушение развития аллантоиса может приводить к аномалиям этого органа. ПУПОЧНЫЙ КАНАТИК. Главным источником развития пупочного канатика является мезенхима амниотической ножки, а также желточного мешка (стебелька). В пупочный канатик включаются аллантоис и растущие по нему сосуды. После образования туловищных складок пупочный канатик оказывается покрытым с поверхности амниотической оболочкой. В последующем желточный мешок и аллантоис постепенно редуцируются. В пупочном канатике (рис. 6.3) проходят две пупочные артерии и одна пупочная вена. Основу его составляет слизистая (студенистая) ткань (вар-тонов студень), относящаяся к соединительным тканям со специальными свойствами. В основном веществе этой ткани содержится большое количество гиалуроновой кислоты, обладающей гидрофильными свойствами. Из-за аккумуляции большого количества воды студенистая ткань имеет выраженные упругие свойства, что препятствует ее сжатию. Снаружи пупоч-ный канатик покрыт амниоти-ческой оболочкой, которая срастается со студенистой тканью.  ФУНКЦИИ. Связь эмбриона с плацентой и проведение из нее к телу эмбриона кровеносных сосудов. При этом студенистая ткань препятствует Г пережатию кровеносных сосудов пупочного канатика при меха-| нических воздействиях и, кроме [ того, участвует в защитной ' функции, т.к. препятствует проникновению из плаценты к эмбриону внесосудистым путем повреждающих веществ. ХОРИОН. Часть незародышевой мезенхимы, которая заполняет полость зародыша, подходит к трофобласту и вступает с ним в тесный контакт. Трофобласт вместе с этой мезенхимой образует третий провизорный орган — хорион (см. рис. 5.10, 6.2). В дальнейшем хорион претерпевает ряд изменений. В его развитии выделяют три периода. 1. Предворсинчатый период (7—8-й день развития). 2. Период образования первичных, вторичных и третичных ворсин (9—50-й дни эмбриогенеза). 3. Период формирования котиледонов (50—90-й дни развития). В предворсинчатый период на поверхности хориона практически отсутствуют выпячивания. Во второй период происходит образование ворсин. Внача-ле первая генерация симпластотрофобласта хориона разрушается, а клетки цитотрофобласта в отдельных участках делятся и образуют выпячивания На поверхности этих выпячиваний образуется вторая генерация симплас тотрофобласта. Так формируются первичные ворсины (9—10-й дни). Затем в эти ворсины прорастает внезародышевая мезенхима и формируются вторичные ворсины (11 — 13 дни). Наконец, после образования первичных крове поеных сосудов в желточном .мешке и образования аллантоиса, сосуды по аллонтоису доходят до вторичных ворсин и врастают в них. Так формируются третичные ворсины (3-я неделя эмбриогенеза). Указанные изменения происходят с той частью хориона, которая обраще на в сторону стенки матки. Он называется ворсинчатым хорионом (ch. frondosum). Напротив, та часть хориона, которая обращена в сторону полости матки, теряет ворсины. Это гладкий хорион ("лысый", "голый" хорион, ch. laeve) (рис. 6.2 г). Ворсинчатый хорион разрушает сосуды слизистой матки, проникает внутрь этих сосудов, и его ворсины омываются кровью. ФУНКЦИЯ хориона — образование плаценты. |