МЯДЕЛЕЦ ОБЩ.ГИСТ.. Литература для медицинских вузов все факультеты о. Д. Мяделец основы цитологии, эмбриологии и общей гистологии
 Скачать 4.75 Mb. Скачать 4.75 Mb.
|

|
Глава 12 МЫШЕЧНЫЕ ТКАНИ Общая морфофункционалытя характеристика. Мышечные ткани пр« ставляют собой тип тканей, объединенных общей функцией — функцт сократимости. Скелетная мышечная ткань обеспечивает передвижение те.; в пространстве. Гладкая мышечная ткань приводит в движение степ к: внутренних органов и сосудов. Сердечная мышечная ткань осуществля< движение кропи по сосудам. Мионейральная ткань обеспечивает изменен!;. размеров зрачка, а миоэпителиальная ткань способствует выведению сскре та из желез. Таким образом, двигательные процессы в организме разнооб разиы, но основой их является сокращение миофибрилл — специальны органоидов движения, состоящих из актиновых и миозиновых филаментоь Двигательные процессы, основанные на взаимодействии сократимых бел ков цитоскелета, имеют место в любой клетке организма, однако в мышеч ной ткани они являются основной и практически единственной функцией Структурной основой этой функции являются сформировавшиеся в фило генезе на базе сократимых белков цитоскелета миофибриллы. Следователь но, второе, что объединяет вес виды мышечных тканей — это наличие в цитоплазме специальных органелл движения миофибрилл. 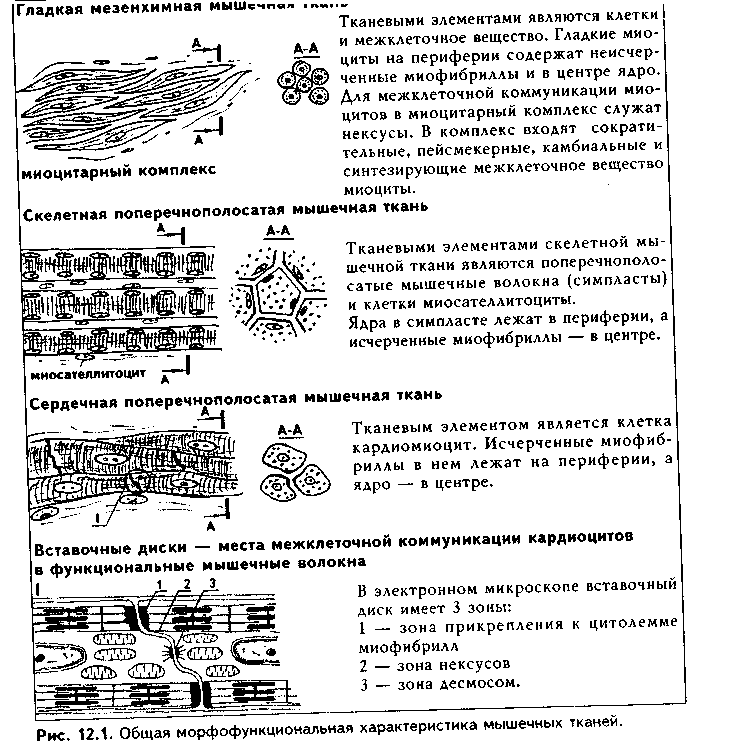 В остальном между пятью указанными выше видами мышечных тканей больше различий, чем сходства. Они имеют разное происхождение. Источником развития скелетной мышечной ткани являются миотомы сомитов. Сердечная мышечная ткань имеет источником развития миоэпикардиальную пластинку, часть висцерального листка спланхнотома. Источник развития гладкой мышечной ткани — мезенхима, в основном спланхнотомная. Мионейральная ткань развивается из нейроэктодермы, а миоэпителиальная — из кожной эктодермы. Тканевыми элементами скелетной мышечной ткани являются симпласты и клетки миосателлитоциты. Все остальные мышечные ткани построены исключительно из клеток: в сердечной мышечной ткани они называются кардиомиоцитами, в гладкой — гладкими миоцитами, в мио-нейралыюй — мионейроцитами, в миоэнителиадыюй — миоэпителиоцитами (или корзинчатыми клетками). Мышечные ткани выполняют своп функции при тесном взаимодействии с нервной тканью. При этом скелетная мышечная ткань получает соматическую двигательную иннервацию, остальные виды мышечной ткани иинервируются вегетативной нервной системой. Регенераторные свойства мышечных тканей различные. Скелетная мы шечная ткань содержит камбиальные клетки (миосателлитоциты) и при необ ходимых условиях регенерирует удовлетворительно, сочетая клеточную и внутриклеточную регенерацию. Содержат камбий и хорошо регенерируют на клеточном уровне гладкая мышечная ткань и миоэпителиальиая ткань. В сер дечной мышечной ткани в дефинитивном состоянии стволовые клетки отсут ствуют, поэтому у взрослого человека подавляющая часть кардиомиоцитов не делится и при повреждении замещается соединительной тканью. Регенерация этой ткани происходит только на внутриклеточном уровне. Регенераторные свойства мионейралыгой ткани не изучены. Общая морфофункциональная характеристика основных видов мышечных тканей приведена на рис. 12.1. КЛАССИФИКАЦИЯ МЫШЕЧНЫХ ТКАНЕЙ Существует несколько подходов в классификации мышечных тканей. 1. Физиологическая классификация. По этой классификации мышечные ткани делятся на произвольные и непроизвольные. К произвольной мышечной ткани относится скелетная мышечная ткань. Ее сокращение контролируется сознанием. Сокращения у этой ткани быстрые, в сокращенном состоянии мышца может находиться относительно непродолжительное время, расслабление также осуществляется быстро. К непроизвольным мышечным тканям относятся все остальные мышечные ткани. При этом у гладкой мышечной ткани сокращения происходят в течение достаточно длительного времени, в сокращенном состоянии гладкие миоциты могут находиться длительно, а процесс расслабления также длительный. У сердечной мышечной ткани сокращения автоматические. Отличия в сократительных актах объясняются особенностями иннервации мышечных тканей, а также наличием в сердечной мышечной ткани проводящих кардиомиоцитов (см. ниже). 2. Морфологическая классификация. Основана на структурном феномене наличия или отсутствия исчерченности миофибрилл. По этой классификации мышечные ткани делятся на: 1) исчерченные; 2) неисчерченные. К неисчерченным мышечным тканям относятся мышечная ткань ме-зенхимного происхождения (сосудов и внутренних органов), эктодермаль-ного происхождения (миоэпителиоциты потовых, молочных, слюнных, слезных желез), мионейрального происхождения (мышцы суживающие и расширяющие зрачок). Компоненты миофибрилл в этих тканях расположены так, что отсутствует их исчерченность. К исчерченным мышечным тканям относят скелетную и сердечную мышечную ткани. В них миофиб-риллы построены из структурно-функциональных единиц саркомеров (см. ниже), в которых имеются светлые и темные участки, придающие миофиб-рилле и всему мышечному волокну исчерченность. 3. Гистогенетическая классификация мышечных тканей учитывает источники их развития. Эта классификация выглядит так: 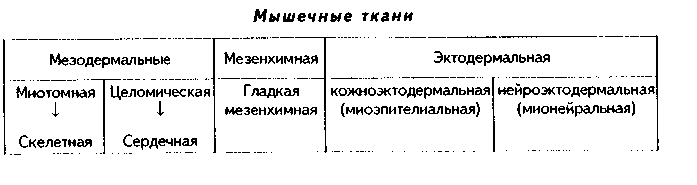 ХАРАКТЕРИСТИКА РАЗНОВИДНОСТЕЙ МЫШЕЧНЫХ ТКАНЕЙ. ГЛАДКАЯ МЫШЕЧНАЯ ТКАНЬ. МЕЗЕНХИМНАЯ ГЛАДКАЯ МЫШЕЧНАЯ ТКАНЬ ГИСТОГЕНЕЗ. Источником развития гладкой мышечной ткани является н основном спланхнотомная мезенхима. Ее клетки мигрируют и окружают эпителиальные зачатки тех органов, в состав которых входит гладкая мышечная ткань. Начало дифферсицировки гладкой мышечной ткани характеризуется удлинением мезенхимных клеток и превращением их из звездчатых в веретеновидные. В цитоплазме клеток появляются органеллы белкового синтеза, осуществляющие синтез специфических белков миофибрилл. Из этих белков производится сборка большого количества миофибрилл, и клетки начинают реагировать на раздражение сокращением. Часть клеток остается в малодифферснцированном состоянии и служит источником для регенерации.  СТРОЕНИЕ. Во взрослом организме гладкая мышечная ткань входит в состав стенки органов пищеварительного тракта, образует мышечные оболочки кровеносных и лимфатических сосудов, бронхиального дерева, яйцеводов, матки, мочеточников, мочевого пузыря, входит в состав капсулы селезенки, есть в эндокарде. Гладкая мышечная ткань стенки сосудов по ряду морфофункциональных признаков отличается от гладкой мышечной ткани другой локализации. Структурно-функциональным тканевым элементом ткани является гладкий миоцит (иногда в качестве второго тканевого элемента называют межклеточное вещество, которое способны синтезировать миоциты). Гладкий миоцит (рис. 12.2) — клетка веретеновидной формы, длина может быть от 20 до 500 мкм, как, например, в матке, в мышечной оболочке которой миоциты имеют также особую (звездчатую) форму. Ядра клеток палочковидной или эллипсоидной формы, с плотным хроматином и 1—2 ядрышками. Гладкий миоцит покрыт цитолеммой. Снаружи от нее лежит тонкая базальная мембрана с ретикулярными фибриллами, которая отграничивает каждый гладкий миоцит от соседних миоцитов. В цитоплазме гладких миоцитов имеются все органеллы общего назначения, лежащие в околоядерных участках цитоплазмы (рис. 12.3): гранулярная ЭПС, выполняющая синтез белков межклеточного вещества, комплекс Гольджи, митохондрии, многочисленные пузырьки, лежащие под цитолеммой (кавеолы) и открытые в сторону межклеточной среды. Этот везикулярный аппарат депонирует ионы Са2*, необходимые для сокращения, и является аналогом одновременно и саркоплазматического ретикулу-ма (СПР), и Т-трубочек в исчерченной мышечной ткани (см. ниже). В ми-оците имеются также и элементы редуцированного саркоплазматического ретикулума в виде пузырьков и небольших цистерн. С ними кавеолы мо- гут иметь связи. Кавеолы СИР содержат в своей мембр.; не белки транспорта кальки: В цитоплазме гладких миоцп тов есть включения гликоген;! 11а периферии миоцитоь под их цитолеммой находят; плотные тельца, состоящие и белка а-актинина — аналоги / линий саркомеров (см. ниже) Есть две разновидности плот ных телец: 1) связанные i внутренней поверхностью илазмолеммы (сарколеммы; миоцита при помощи комплекса адгезивных белков (винкули-на, тензина и др.). Эти плот ные тельца на самом деле представляют собой срезанные поперечно пластинки, имеющие вид длинных непрерывных ребер, которые лежат параллельно друг другу под сарколеммой; 2) свободно лежащие в цитоплазме (саркоплазме). Лежат в виде правильной цепочки. К плотным тельцам прикрепляются актиновые и промежуточные десминовые филаменты. Последние образуют сложную трехмерную сеть в саркоплазме. Важный компонент цитоплазмы гладких миоцитов — сократительные белковые нити, или миофиламенты, образующие миофибриллы. Эти нити расположены вдоль длинной оси миоцита, а по отношению друг к другу так, что не образуют поперечной исчерченности. Тонкие актиновые миофиламенты одним концом прикрепляются к плотным тельцам. Они в отличие от скелетной мышечной ткани состоят только из белка актина (мышечного и немышечного), не содержат тропонина и тропомиозина и более многочисленны. Актиновые филаменты взаимодействуют с толстыми мио-зиновыми филаментами, образуя так называемые сократимые единицы. В отличие от миозиновых филамептов скелетной мышечной ткани миозино-вые филаменты гладких миоцитов менее стабильны, а по мнению некоторых исследователей, молекулы миозина в состоянии покоя находятся в де-полимеризованной форме, и миозиновые филаменты организуются путем сборки непосредственно перед сокращением, вновь распадаясь после него. Поэтому в гладких мышечных тканях не формируются миофибриллы, сар- 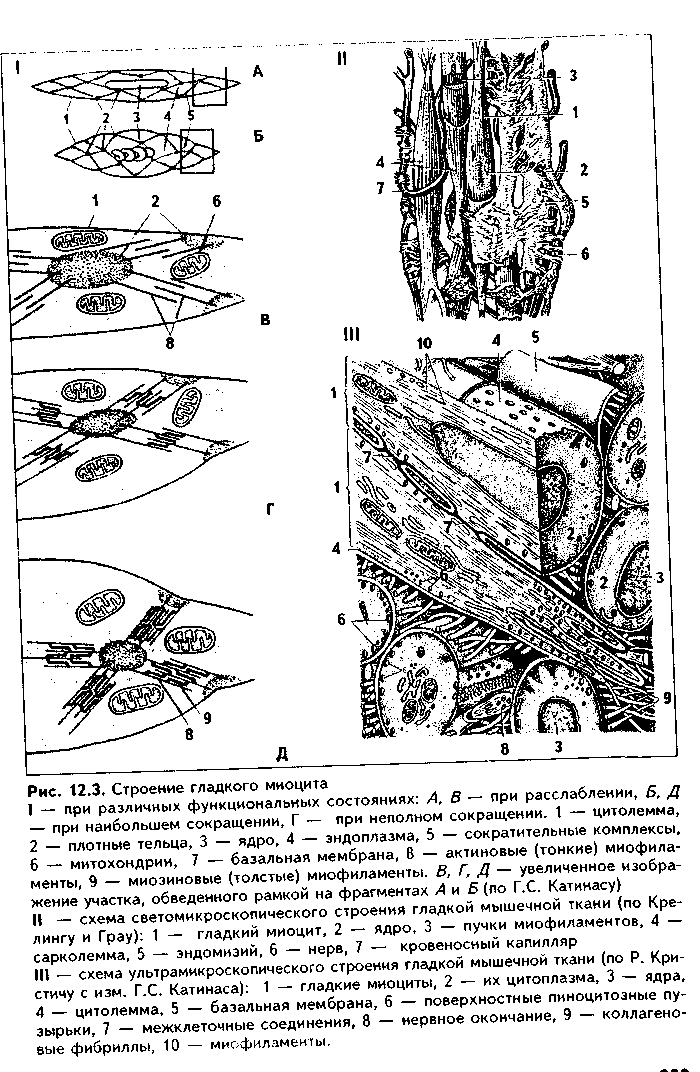 комсры и отсутствует поперечная исчерченность. Сборка из молекул мио зипа толстых миозииовых филаментов происходит при инициации сокрп тения, и этот процесс, а также взаимодействие актиновых и миозиновьп филаментов активируют ионы кальция, поступающие из кальциевых депо -- СПР, кавеол и митохондрий. Образующиеся сократительные единицы направлены под углом к длине миоцита. Механизм сокращения гладких миоцитов принципиально сходен с со кращением скелетных мышечных волокон и более подробно будет рассмотрен ниже. Он заключается во взаимодействии актиновых и миозииовых филаментов (теория скольжения X. Хаксли), которое инициируют ионы кальция, выделяемые СПР, митохондриями и кавеолами. Под действием нервного импульса из пиноцитозных пузырьков высвобождается Са2', который образует комплекс с кальцийсвязывающим белком кальмодулином. Комплекс "Са2+ — кальмодулин" активирует фермент киназу легких цепей миозина, фосфорилирующую легкие цепи миозина. Фосфорилирование миозина придает ему способность взаимодействовать с актиновыми фила ментами. В итоге головки молекул миозина сформированных и активиро ванных миозииовых филаментов начинают взаимодействовать с активны ми центрами актиновых филамент, т.к. обладают свойством липкости Они совершают тянущие гребковые движения, скользя вдоль актиновых филамент. В результате повторяющихся гребковых движений миозииовых филамент вдоль актиновых сближаются плотные тельца, и гладкий мио цит сокращается. Промежуточные десминовые филаменты препятствуют сильной деформации клетки при ее сокращении. Для сокращения необходима энергия АТФ, гидролиз которой происходит медленно, что отражается на скорости сокращения. Прекращает сокращение фермент фосфатаза миозина, отщепляющая фосфат от легких цепей миозина (дефосфорилировапие). При этом особенность гладких мыщц заключается в том, что не все миозиновые мостики после дефосфорилирования разрушаются: часть головок миозина остается связанной с актиновыми филаментами. Это обеспечивает длительное поддержание тонуса гладких мышц без дополнительных энергетических затрат. Гладкие миоциты функционируют не изолированно, а формируют ми-оцитарные комплексы. Нервные окончания подходят не ко всем миоцитам. а только к одному в комплексе. Комплекс состоит из 10—12 миоцитов. В составе комплекса миоциты тесно взаимодействуют друг с другом при помощи десмосом и нексусов — щелевых контактов. В области нексусов банальные мембраны миоцитов прерываются. Через нексусы происходит передача возбуждения от одного миоцита к соседним, и в результате сокращением охватывается весь комплекс. В состав миоцитарного комплекса входят несколько различающихся по функции миоцитов. 1. Сократительные миоциты преимущественно вы-полняют сократительные акты. 2. Секреторные миоциты синтезируют и секретируют межклеточное вещество. 3. Миоциты-пейсмекеры генерируют потенциал действия и передают его на соседние клетки. 4. Камбиальные (малодифференцированные) миоциты служат источником регенерации мышечной ткани. Имея мезенхимное происхождение, гладкие миоциты генетически очень близки фибробластам и другим аналогичным клеткам-продуцентам межклеточного вещества: они способны к синтезу собственного межклеточного вещества гладкой мышечной ткани, которое иногда рассматривают как второй тканевой элемент гладкой мышечной ткани. Регенерация гладкой мышечной ткани происходит не только за счет ма-лодиффсРС1щиР01шшых клеток, но и за счет адвентициальных клеток (возможно, за счет перицитов), а при повреждении — за счет миофибробластов в силу их близкого генетического родства. Возможна и внутриклеточная регенерация гладких миоцитов, основанная на восстановлении органелл, их гипертрофии и гиперплазии. МИОЭПИТЕЛИАЛЬНАЯ ТКАНЬ. Тканевым элементом этой ткани является миоэпителиоцит, или корзинчатая клетка. Источником развития этой ткани является кожная эктодерма. Органная локализация — концевые отделы и некоторые выводные протоки потовых, молочных, слезных, слюнных желез. Эти клетки дифференцируются из эктодермы одновременно с секреторными клетками. При этом миоэпителиоциты плотно прилегают к экзокриноцитам концевых отделов. Строение. Миоэпителиоциты имеют звездчатую форму и своими отростками окружают концевые отделы (рис. 12.4). В отростках есть актиновые филаменты, сборка миозииовых филаментов происходит накануне сокращения и активируется ионами Са2+, вышедшими из кальциевого депо под воздействием нервного импульса. В результате этого миофибриллы не имеют поперечной исчерчеиности. Сокращение отростков ведет к сдавле-нию концевого отдела и выведению из него секрета. Снаружи от миоэни-телиоцитов находится базальная мембрана. Регенерация. Среди дифференцированных миоэпителиоцитов имеются менее дифференцированные клетки, обладающие признаками камбиальных. За счет их митотического деления и дифференцировки в сократимые миоэпителиоциты происходит регенерация миоэпителиалыюй ткани. По другим сведениям, регенерация этой ткани происходит за счет камбиальных клеток многослойного эпителия, дифференцирующихся как в секреторные, так и в миоэпителиальные клетки. МИОНЕЙРАЛЬНАЯ ТКАНЬ. Эта разновидность мышечных тканей входит в состав мышц радужной оболочки глаза — мышцы суживающей и мышцы расширяющей зрачок. Источником развития мионейральной ткани является нейроэктодерма. Структурно-функциональным элементом ми-онейральной ткани является мионейроцит, или миопигментоцит. Это одноядерные веретеновидные клетки. Содержат в цитоплазме гладкие миофиб-риллы, которые состоят из тонких актиновых миофиламснтов, расположенных так же, как в гладкой мезенхимной мышечной ткани. Толстые ми-озиновые филаменты формируются при инициации сокращения ионами Са2+. В клетках много митохондрий и пигментных гранул. Между клетками есть нексусы и десмосомы. Иннервация мионейральной, так же как и гладкой мышечной ткани, выполняется за счет вегетативной нервной системы. Регенераторные свойства этой ткани не изучены.  К видоизмененным гладким миоцитам относятся также эндокринные миоциты мышечной оболочки приносящих и выносящих артериол почечных телец, секретирующие гормон ренин (юкстагломерулярные клетки). Характеризуются сильно развитым белоксинтезирующим аппаратом и редуцированным сократительным аппаратом. Следует также упомянуть, что выраженной сократительной функцией обладают видоизмененные фиб-робласты — миофибробласты. СКЕЛЕТНАЯ МЫШЕЧНАЯ ТКАНЬ Функции. 1) Функция движения. Входит в качестве основной ткани в состав скелетных мышц — органов движения. Обеспечивает перемещение тела в пространстве и частей тела друг относительно друга. Кроме скелетных мышц, входит в состав мышц языка, стенок полости рта, пищевода, гортани, анального отдела прямой кишки, образует глазодвигательные мышцы. 2) Терморегуляционная функция. Сокращение скелетной мышечной ткани ведет к образованию большого количества тепла, что в условиях холода обеспечивает согревание тела (так называемый сократительный термогенез). 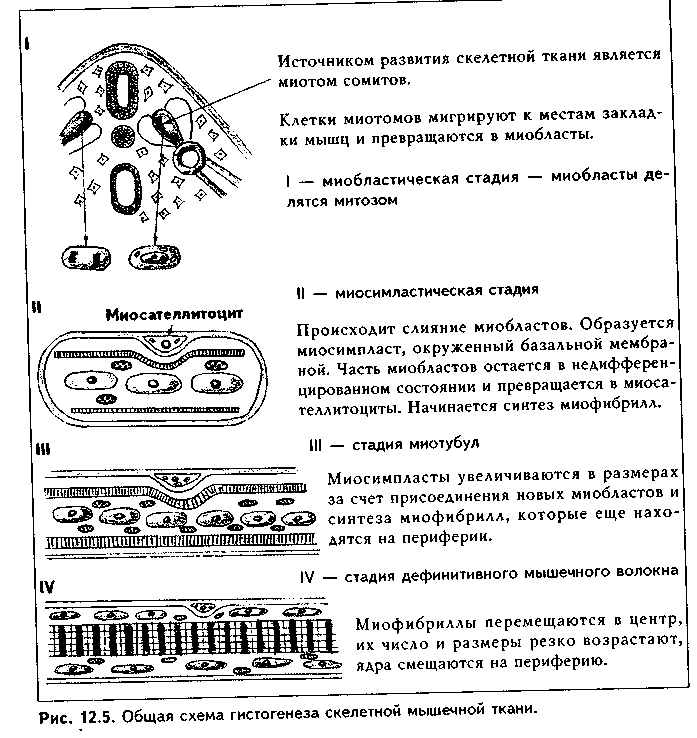 Тканевыми элементами скелетной мышечной ткани являются как сим-пласты (поперечнополосатые мышечные волокна), так и клетки — миоса- теллитоциты. ГИСТОГЕНЕЗ СКЕЛЕТНОЙ МЫШЕЧНОЙ ТКАНИ. Источником ее развития являются миотомы сомитов. Основная масса клеток миотомов превращается в митотические миобласты (С,-миобласты), которые делятся митозом (пролиферативный митоз) (рис. 12.5). Часть С,-миобластов при этом обособляется в виде миосателлитоцитов. Остальные миобласты пр.. должают делиться при помощи квантального (дифференцирующего) ми п. за, и, дифференцируясь, превращаются в ностмитотические миоблас i ■ (Gd-миобласты) — одноядерные веретеновидиые клетки, приобретают! i способность к синтезу специфических белков. Они мигрируют из миотомо, в места закладки будущих мышц. Среди мигрирующих миобластов нам. дятся и миосателлитоциты, сохраняющие свойства малодифференцироваи пых клеток до конца жизни. Эта стадия называется миобластической (I ста дия миогенеза). |