МЯДЕЛЕЦ ОБЩ.ГИСТ.. Литература для медицинских вузов все факультеты о. Д. Мяделец основы цитологии, эмбриологии и общей гистологии
 Скачать 4.75 Mb. Скачать 4.75 Mb.
|

|
Часть мезенхимы развивается из нейроэктодермы ганглиозных пластинок — эктомезенхима. Является источником развития хрящевых и костных тканей черепа, соединительной ткани пульпы зуба, дентинобластов, соединительной ткани головы. Часть мезенхимы образуется из энтодермы — энтомезенхима. Мезенхимные клетки обладают выраженной подвижностью, в процессе образования мезенхимы мигрируют между зародышевыми листками и заполняют все пространство между ними. Строение. Мезенхимные клетки отросчатые, соединяются друг с другом отростками в "ложный" синцитий. Между клетками есть слизистое межклеточное вещество из тканевой жидкости и тонких фибрилл. Функции. 1. Роль эмбриональной соединительной ткани — защитная, трофическая, опорная, регуляторная функция в теле зародыша. 2. Мезенхима является эмбриональным зачатком, из которого образуются ткани внутренней среды, или соединительные ткани в широком смысле, а также гладкая мышечная ткань. Общая морфофункциональная характеристика тканей внутренней среды 1. Общий источник развития — мезенхима. 2. Все эти ткани состоят из двух типов тканевых элементов — клеток и межклеточного вещества. 3. Клетки анолярны (не имеют полюсов). 4. Ткани в подавляющем большинстве полидифферонпые. 5. В подавляющем большинстве случаев богато васкуляризованы (существует афоризм: "нет соединительной ткани без сосудов и нет сосудов без соединительной ткани"). 6. Как правило, все ткани мезенхимного происхождения хорошо регенерируют, т.к. это камбиальные обновляющиеся ткани. 7. Ткани мезенхимного происхождения выполняют похожие функции: барьерно-защитную, трофическую, опорную, регуляторную, пластическую (участие в воспалении, регенерации, компенсаторно-приспособительных реакциях). КРОВЬ И ЛИМФА Кровь и лимфа, как и все образующиеся из мезенхимы ткани, состоят из клеток и межклеточного вещества. Клетки крови называются форменными элементами. Межклеточное вещество представлено плазмой, находится в жидком состоянии. Соотношение форменных элементов и плазмы равно 40:60, это соотношение называется гематокритом и в клинике является показателем степени сгущения или разжижения крови. Кровь входит в состав так называемой функциональной системы крови, которая помимо самой крови включает органы кроветворения и крове-разрушения.  ФУНКЦИИ КРОВИ 1. Транспортная функция. Включает в себя целый ряд частных функций: а) трофическая функция заключается в транспорте питательных веществ из мест всасывания и накопления к клеткам и тканям; б) дыхательная функция — перенос кислорода из легких к клеткам и тканям и углекислого газа от тканей к легким; в) экскреторная функция заключается в переносе конечных продуктов из тканей к органам выделения (кожа, почки) и участии в их выведении из организма с потом и мочой (пот и моча являются своеобразными фильтратами плазмы крови); г) регуляторная функция состоит в том, что с кровью транспортируются гормоны, медиаторы, цитокины и другие биологически активные вещества, регулирующие функции клеток, тканей и органов; д) терморегуляционная функция зак-лючается в переносе тепла, его распределении между органами и выделешн во внешнюю среду. Кровью транспортируются также микроорганизмы и и токсины. 2. Защитная функция. Клетки крови участвуют в иммунных и воспалительных реакциях. 3. Гомеостатическая функция основывается на рассмотренных преды дущих функциях и заключается в сохранении и поддержании постоя нети.. внутренней среды: метаболического, энергетического, кислотно-щелочной-осмотического, температурного, антигенного и т.д. гомеостаза. СТРОЕНИЕ КРОВИ Плазма. Это своеобразное межклеточное вещество жидкой консистеп ции Состав плазмы такой: 90% воды; 9% органических веществ (белки альбумины, глобулины, фибриноген, компоненты комплемента; линиды углеводы); 1% минеральных веществ. Белки плазмы крови продуцируются печенью за исключением у-глобулипов (образуются плазмоцитами) Белки крови создают ее вязкость, онкотическое давление, обеспечивают коагуляцию крови и лимфы, выполняют защитные и транспортные функции При коагуляции крови фибриноген переходит в нерастворимый белок фибрин, оставшаяся жидкая часть плазмы называется сывороткой крови. В сыворотке находятся антитела, поэтому донорская сыворотка очень часто используется с лечебной целью. Форменные элементы крови подразделяются на три вида: эритроциты лейкоциты, тромбоциты. Для изучения форменных элементов крови используют мазки, окрашенные азур-2-эозином или другими красителями. 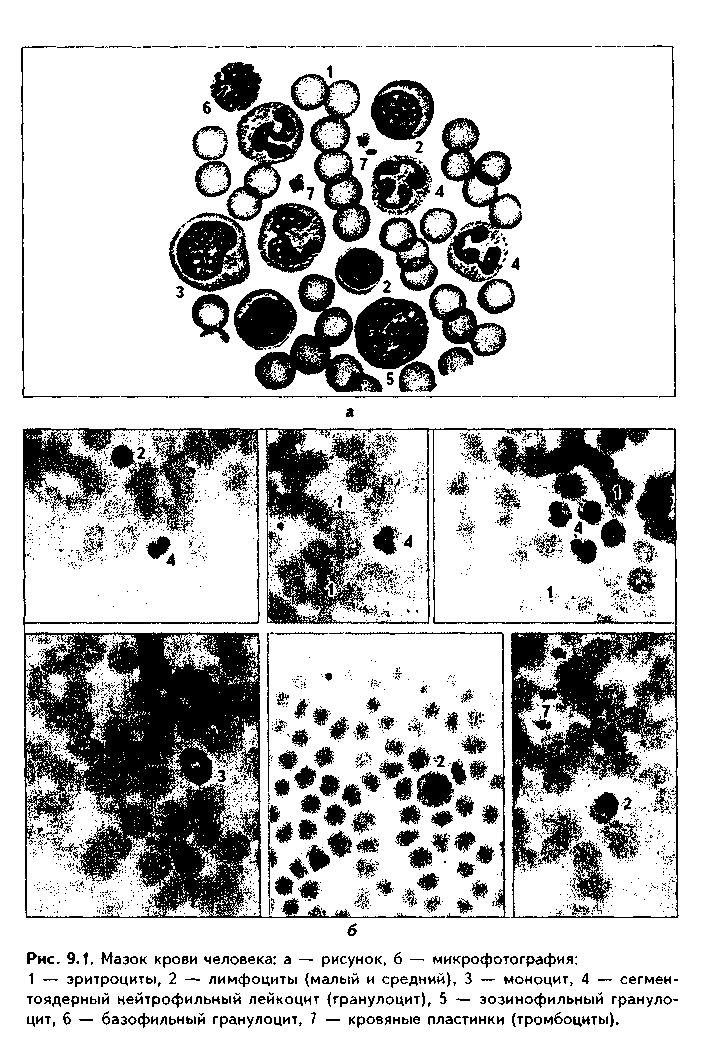 ЭРИТРОЦИТЫ. Это безъядерные красные кровяные клетки постклеточные структуры (рис. 9.1). Большинство эритроцитов имеет форму двояковогнутого диска (дискоциты), которые преобладают в крови. Эта форма характерна для молодых и зрелых полноценных эритроцитов. Благодаря дисковидной форме по сравнению со сферической происходит увеличение поверхности эритроцита в 1,5 раза. Одновременно существенно уменьшается диффузионное расстояние для газов, создаются возможности, во-первых, для увеличения размера эритроцитов без их разрыва (до определенных пределов) в гипотонической среде, а во-вторых, для обратимой деформации при прохождении через узкие капилляры. Дисковидная форма эритроцитов поддерживается благодаря деятельности осмотических насосов, создающих определенный уровень осмотического давления, и цитоске-лста. Могут встречаться также сферические эритроциты - сфероциты, эритроциты с зазубренными краями - эхиноциты, серповидные эритроциты, дрепаноциты (каплевидные эритроциты), стоматоциты (куполообразной формы), планоциты (плоские эритроциты) и др. Сферическая форма харак-терна для стареющих эритроцитов, а также при врожденном сфероци-тозе. Такие эритроциты неустойчивы к деформации, колебаниям осмотического давления, другим воздействиям и подвергаются массивному разрушению. Серповидные эритроциты наблюдаются при серповидноклеточ-ной анемии, обусловленной аномалией гемоглобина (в результате мутации гена нормальный гемоглобин (НЬА) заменяется на патологический гемоглобин S (HbS)). Серповидные эритроциты также нестойки и имеют малую продолжительность жизни. Изменение формы эритроцитов называется пойкилоцитозом. 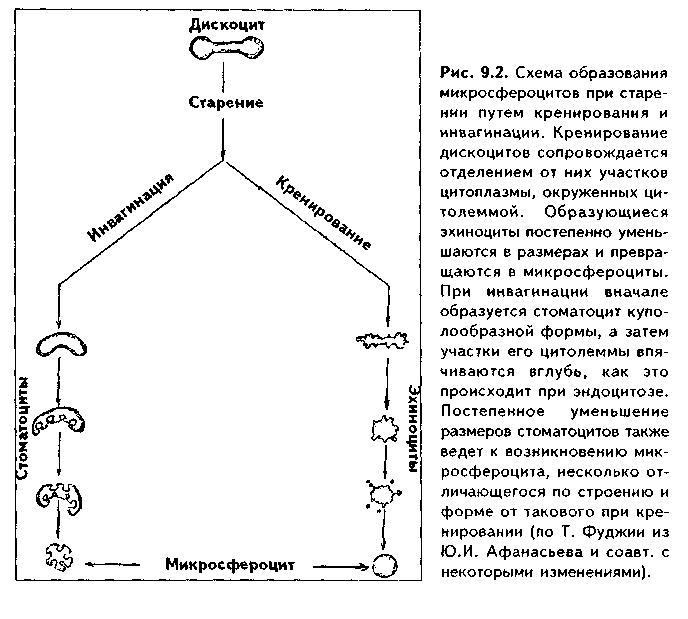 При старении эритроцитов они превращаются в микросфероциты. Это превращение может происходить двумя путями: путем кренирования и путем инвагинации (рис. 9.2). Эритроциты имеют диаметр 7—8 мкм и толщину 2 мкм. Такие эритроциты называются нормоцитами. В крови содержится также небольшое количество макроцитов (с диаметром до 10 мкм), гигантоцитов (диаметр 12 и более мкм), микроциты (с диаметром 6 мкм). Появление в крови эритроцитов различной величины называется анизоцитозом.  Количество эритроцитов в крови у мужчин составляет 4,5—5,3х10,2/л, у женщин — 4—4,5x10,2/л. Снижение числа эритроцитов называется эритро-пенией, или малокровием, тогда как увеличение — эритроцитозом. Электронномикроскопическое строение эритроцитов показано на рис. 9.3. Эритроцит ограничен плазмолеммой толщиной 20 им, которая обладает избирательной проницаемостью для веществ. Снаружи плазмолемма покрыта гликокаликсом, который содержит антигены А и В, определяющие группы крови. Под плазмолеммой эритроцита находятся компоненты цитоскелета. Он образован двумерной гибкой сетью филаментов, построенных из белка спектрина. Спектрин формирует две закрученные наподобие веревки цепи (а и (3). Две цепи связаны между собой при помощи актина и белка полосы 4.1. Это так называемые узлы. В свою очередь, узлы при помощи белка анкирина "пришиты" к трансмембранному ре-цепторному белку полосы 3, который может прикрепляться к трансмембранному белку-рецептору гликофорину. Белок полосы 3 и гликофорин являются гликопротеинами, углеводные цепи которых формируют основную часть гликока-ликса, причем только гликофорин содержит антигенные детерминанты — агглютиногены системы А, В, 0 (рис. 9.4). Благодаря цитоскелету эритроцит способен к значительной деформации. При этом в состоянии покоя спектрииовые цепи закручены равномерно, при деформации они раскручиваются в одних участках и еще сильнее закручиваются в других. Это ведет к обратимому изменению формы эритроцита. Однако при резкой деформации связи элементов цитоскелета между собой и с плазмолеммой могут разорваться, и тогда эритроцит теряет способность к возвращению к первоначальной форме. Дефекты белков цитоскелета лежат в основе их повышенного разрушения при старении, а также ряде заболеваниях (сфероцитозе, сериовидноклеточной анемии и др.). Основную массу эритроцита составляет гемоглобин. При электронной микроскопии он выявляется в виде очень плотных гранул размером 4—5 пм, а в световом микроскопе обеспечивает оксифилию цитоплазмы. Кроме гемоглобина в цитоплазме находятся до 60% воды, глюкоза (основной источник энергии), АТФ и ферменты, в основном гликолитическис и ненто-зофосфатного пути. Могут также встречаться единичные мелкие мембранные пузырьки. Другие органеллы в эритроците отсутствуют, он утрачивает их в процессе дифферепцировки. Гемоглобин — дыхательный пигмент. Состоит из белка глобина и железосодержащей части — гема. Гем легко присоединяет кислород, в результате гемоглобин превращается в оксигемоглобин. Это происходит в капиллярах легкого, где высокое парциальное давление кислорода (100 мм рт.ст.). В капиллярах органов и тканей давление кислорода меньше — 40 мм рт.ст., поэтому там происходит диссоциация оксиге-моглобина на кислород и гемоглобин. Кислород легко поступает в ткани, потому что в них давление кислорода еще меньше — 20 мм рт.ст. Углекислый газ также может транспортироваться в связанной с гемоглобином форме, но большая его часть в эритроците связывается с водой с образованием углекислоты. В легких она расщепляется до воды и углекислого газа, который выделяется в плазму, а затем в выдыхаемый воздух. В гипотонической среде эритроциты накапливают воду и разрушаются (гемолиз). В гипертонической среде они, наоборот, отдают воду и сморщиваются (плазмолиз). Наряду со зрелыми эритроцитами в крови могут быть незрелые эритроциты — ретикулоциты. Они имеют сферическую форму, в их цитоплазме при специальной окраске крезиловым или метиловым синим выявляется сеть — ретикулум. Это остатки органелл: небольшое число свободных рибосом, митохондрий, центриоль, элементы комплекса Гольджи (благодаря наличию сети клетки и названы ретикулонитами). Из-за сферической формы ретикулоциты в функциональном отношении много хуже, чем зрелые эритроциты, способны транспортировать газы. При созревании ретикулоцитов в них завершается оформление цитоскелета, утрачиваются остатки органелл и ряд рецепторов, возрастает содержание гемоглобина. Т.к. ретикулоциты при обычной окраске не отличаются от зрелых эритроцитов, то определение их числа специальным окрашиванием имеет большое значение для выявления скрытой анемии. В норме число ретикулоцитов равно 1—2% от всех эрит]юцитов, повышено у новорожденных (до 6—7%) т! детей первого года жизни. Их количество возрастает также при кровотечении, массивном гемолизе и при подъеме на высоту.  Время жизни эритроцитов в крови составляет 100—120 суток, после чего они разрушаются в селезенке, печени или красном костном мозге. При этом г костном мозге железо захватывается особым видом макрофагов (клетками кормилками), которые передают его вновь образующимся эритроцитам. Функции эритроцитов. 1. Дыхательная — перенос кислорода в ткани и углекислого газа от тканей в легкие. 2. Регуляторная и защитная функции — перенос на поверхности различных биологически активных, токсических веществ, защитных факторов: аминокислот, токсинов, антигенов, ан тител и др. На поверхности эритроцитов часто может происходить реакция антиген-антитело, поэтому они пассивно участвуют в защитных реакциях. Тромбоциты (рис. 9.5, 9.6) представляют собой свободно циркулирующие в крови безъядерные фрагменты цитоплазм:,, гигантских клеток красного ко стного мозга — мегакариоци-тов. Размер тромбоцитов 2—3 мкм, их количество в крови составляет 200-300x109/л. Каждая пластинка в световом микроскопе состоит из двух частей: хромомера, или грануломера (интенсивно окрашенная часть). и гиаломера (прозрачная часть) Хромомер находится в центре тромбоцита и содержит гранулы, остатки органел (митохондрии, ЭПС), а также включения гликогена. Гранулы делятся на четыре вида. 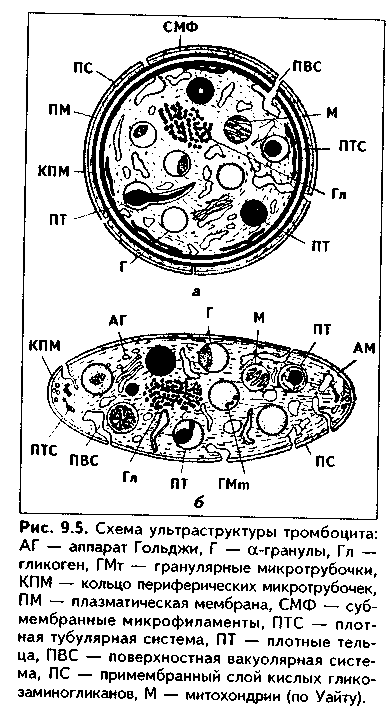 1. а-гранулы содержат фибриноген, фибронектин, ряд факторов свертывания крови, ростовые факторы, тромбоспондин (аналог актомиозинового комплекса, участвует в адгезии и агрегации тромбоцитов) и другие белки. Окрашиваются азуром, давая базофилию цэануломера. 2. Второй тип гранул называется плотными тельцами, пли 8-гра-нулами. Они содержат серотонин, гистамин (поступающие в тромбоциты из плазмы), АТФ, АДФ, кальций, фосфор. АДФ вызывает агрегацию тромбоцитов при повреждении стенки сосуда и кровотечении. Серотонин стимулирует сокращение стенки поврежденного кровеносного сосуда, а также вначале активирует, а затем ингибирует аггрегацию тромбоцитов. 3. ^.-гранулы — типичные лизосомы. Их ферменты выбрасываются при ранении сосуда и разрушают остатки неразрушенных клеток для лучшего прикрепления тромба, а также участвуют в растворении последнего. 4. Микропероксисомы содержат пероксидазу. Их количество невелико. Кроме гранул в тромбоците есть две системы канальцев: 1) канальцы, связанные с поверхностью клеток. Эти канальцы участвуют в экзо-цитозе гранул и эндоцитозе. 2) система плотных трубочек. Образуется за счет деятельности комплекса Гольджи мегакариоцита. Лежат либо под цитоскелетом, либо диссоциированно в цитоплазме. Возможно, накапливают кальций и являются аналогом саркоплазматической сети мышечных волокон. 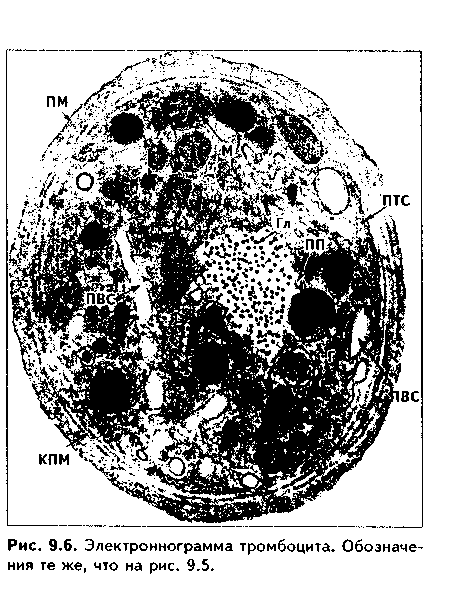 В тромбоцитах есть также цитоскелет, представленный микротрубочками, актиновыми филаментами и промежуточными виментиповыми фи-ламентами. Микротрубочки лежат па периферии и формируют мощный жесткий каркас. Актиповые филамепты, с одной стороны, пронизывают всю цитоплазму, с другой — формируют периферическое сгущение между микротрубочками. Здесь же концентрируются промежуточные филамепты. При образовании тромба в тромбоцитах также быстро образуются и мио-зиновые филамепты, которые взаимодействуют с актиновыми филаментами и вызывают сжатие (ретракцию) тромба. На поверхности тромбоцитов имеется сильно развитый гликокаликс с большим содержанием рецепторов к различным активаторам и факторам свертывания крови. Функции тромбоцитов. 1. Участвуют в свертывании крови и.остановке кровотечения. Активацию тромбоцитов вызывают АДФ, выделяемая поврежденной сосудистой стенкой, а также адреналин, коллаген и ряд медиаторов гранулоцитов, эн-дотелиоцитов, моноцитов, тучных клеток. В результате адгезии и агрегации тромбоцитов при образовании тромба на их поверхности образуются отростки, которыми они слипаются друг с другом. Образуется белый тромб. Далее тромбоциты выделяют факторы, которые превращают протромбин в тромбин, под влиянием тромбина происходит превращение фибриногена в фибрин. В результате вокруг тромбоцитарных конгломератов образуются нити фибрина, составляющие основу тромба. В нитях фибрина задерживаются эритроциты. Так формируется красный тромб. Серото-нин тромбоцитов стимулирует сокращение сосуда. Кроме того, за счет сократимого белка тромбостенина, который стимулирует взаимодействие ак-типовых и миозиновых филаментов, тромбоциты тесно сближаются, тяга передается также на нити фибрина, тромб уменьшается в размерах и становится непроницаемым для крови (ретракция тромба). Все это способствует остановке кровотечения. 2. Тромбоциты одновременно с образованием тромба стимулируют регенерацию поврежденных тканей. 3. Обеспечение нормального функционирования сосудистой стенки, в первую очередь, сосудистого эндотелия. В крови есть пять видов тромбоцитов: а) юные; б) зрелые; в) старые; г) дегенеративные; д) гигантские. Они различаются по строению. Продолжительность жизни тромбоцитов равна 5—10 суткам. После этого они фагоцитируются макрофагами (в основном в селезенке и легких). В крови в норме циркулирует 2/3 всех тромбоцитов, остальные депонированы в красной пульпе селезенки. В норме некоторое количество тромбоцитов может выходить в ткани (тканевые тромбоциты). Нарушение функции тромбоцитов может проявляться как в гинокоа-гуляции, так и в гиперкоагуляции крови. В первом случае это ведет к повышенной кровоточивости и наблюдается при тромбоцитопении и тромбоци-топатии. Гиперкоагуляция проявляется тромбозами — закрытием просвета сосудов в органах тромбами, что приводит к некрозу и гибели части органа. ЛЕЙКОЦИТЫ Лейкоциты — это белые клетки крови. В крови их содержится 3—8х10!'/л. Они делятся на две группы: зернистые, или гранулоциты, и незернистые, или агранулоциты. Зернистые лейкоциты имеют в цитоплазме окрашенные гранулы и сегментированные ядра. В зависимости от окрашивания гранул различают эозинофильные (синоним окси-, ацидофильные), нейтрофильные и базо-фильные лейкоциты. Эозинофильные лейкоциты содержат в основном гранулы, окрашивающиеся кислыми красителями в красный цвет. Нейтрофильные гранулоциты имеют и оксифильные, и базофильные, или азуро-фильные гранулы. Базофильные лейкоциты содержат в цитоплазме гранулы, воспринимающие основные красители. Нейтрофильные лейкоциты в периферической крови могут иметь разную степень зрелости и различные формы ядра. В зависимости от этого есть юные (бобовидное ядро), палоч-коядерные (ядро в виде изогнутой палочки), сегментоядерные (ядра имеют 4—5 сегментов) нейтрофильные лейкоциты. Основная функция гранулоцитов — участие в защитных реакциях организма, фагоцитоз. Незернистые лейкоциты (агранулоциты) делятся на две группы: лимфоциты и моноциты. Их функцией является также защитная, участие в иммунных реакциях. Нейтрофильные лейкоциты. Это наиболее распространенный вид лейкоцитов. Их содержание в крови равно 60—75%. Это округлые клетки размером около 10 мкм (рис. 9.1). В мазке, однако, нейтрофилы распластываются, что ведет к увеличению их размеров до 12-15 мкм. Способны к амебовидному движению, поэтому их форма может меняться. Плазмолемма нейтрофилов содержит развитый гликокаликс с рецепторами ко многим медиаторам, гормонам, цитокинам и другим биологически активным веществам. Цитоплазма клеток слабо оксифильна. В ней содержатся гранулы трех видов (рис. 9.7). 1. Первичные гранулы. Этот тип гранул нейтрофилов окрашивается основными красителями (например, азуром) и называется азурофильными гранулами. Это самые крупные гранулы (0,4—0,8 мкм). Их количество равно 10—20% от всех гранул. Представляют собой типичные лизосомы. Они содержат гидролитические ферменты, переваривающие бактерии. Кроме того, в гранулах содержатся катионные белки, дефензины и другие белки, осуществляющие внутриклеточную, а при их секреции во внеклеточное пространство — и дистантную гибель микроорганизмов (так называемый нефагоцитарный тип бактерицидной активности). В азурофильных гранулах содержатся также компоненты, разрушающие компоненты межклеточного вещества, в частности, эластаза и протеиназа, расщепляющие эластин. Это позволяет клеткам легко мигрировать в ткани, а также осуществлять деградацию компонентов межклеточного вещества, что имеет большое значение как в норме, так и в патологии. 2. Вторичные, оксифильные гранулы называются иначе специфическими, т.к. они составляют до 80% всех гранул. Имеют размеры до 0,2 мкм и поэтому плохо видны в световом микроскопе. В них содержатся фер- менты: лизоцим, щелочная фосфатаза, коллагеназа, пероксидаза, белок фагоци-тин с бактерицидными свойствами, лактоферрин, кати-онные белки, адгезивные белки. Лизоцим (мурамида-за) расщемляет полисахариды бактериальной стенки, в результате бактерии становятся подверженными осмотическому шоку и разрушаются. Щелочная фосфатаза и пероксидаза разрушают ДНК бактерий, фагоцитам и катионные белки обеспечивают нефагоцитарный тип бактерицидной активности. Лактоферрин связывает железо, необходимое для развития бактерий, что подавляет их размножение. Специфические гранулы содержат также коллагеназу, расщепляющую коллаген межклеточного вещества. 3. Третичные (коллагеназ-ные) гранулы открыты в последнее время. Содержат желати-назу, расщепляющую межклеточное вещество, прежде всего компоненты базалыюй мембраны, а также лизоцим и адгезивные белки. Полагают, что адгезивные белки гранул обеспечивают прикрепление нейт-рофилов к эндотелию сосудов, а желатиназа, расщепляя компоненты базалыюй мембраны, способствует миграции через нее клеток. Следовательно, третичные гранулы обеспечивают миграцию нейтрофилов в ткани. Секреторные пузырьки. Это мембранные структуры, несущие множество адгезионных молекул и рецепторов (их своеобразное депо). При активации нейтрофилов секреторные пузырьки сливаются с плазмолеммой и обеспечивают выход на поверхность клеток дополнительных рецепторов и адгезионных молекул. Это имеет особое значение для инициации миграции через стенку сосудов. 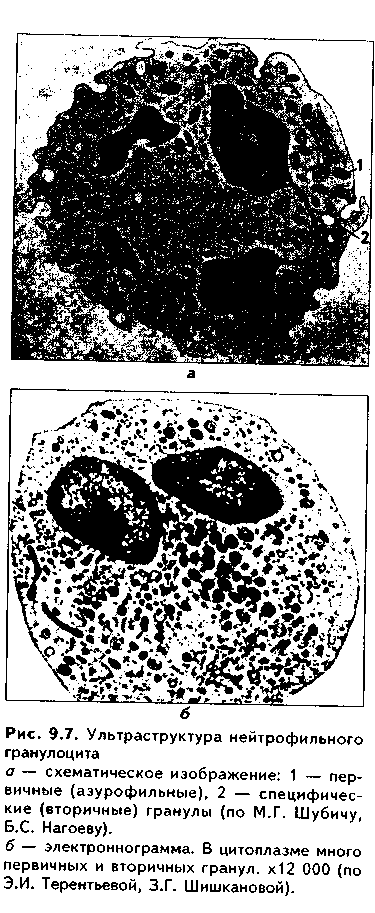 В цитоплазме нейтрофилов кроме гранул находятся митохондрии, ЭПС, комплекс Гольджи, но эти органеллы слабо развиты. Напротив, ци-тоскелет развит хорошо. Ядра нейтрофилов содержат гетерохроматин, поэтому гипербазофильны. Форма ядер зависит от степени зрелости нейтрофилов. Юные нейтрофилы имеют ядро в виде почки или боба. Палочкоя-дерные нейтрофильные лейкоциты содержат ядро в виде изогнутой палочки, сегментоядерные — в виде нескольких сегментов. У женщин ядра нейтрофилов содержат инактивированную вторую Х-хромосому в виде барабанной палочки. При активации нейтрофилов в них происходит так называемый "респираторный взрыв": резкая активация окислительных процессов с образованием активных форм кислорода, губительных для бактерий. При массивной миграции нейтрофилов в очаг воспаления активные формы кислорода (Н202 супероксид-анион и др.), а также ферменты нейтрофилов могут вызывать гибель тканей и самих нейтрофилов с образованием гноя (гнойное воспаление). В норме же нейтрофилы добывают энергию анаэробным путем, что позволяет им функционировать в тканях, обедненных кислородом. Продолжительность жизни нейтрофилов около 8 суток. Свои основные функции они выполняют в тканях, а не в крови (в которой находятся от 1 до 10 ч), поэтому есть тканевый и сосудистый пулы нейтрофилов. Функции. 1. Фагоцитоз. И.И. Мечников назвал их микрофагами. Нейтрофилы фагоцитируют в основном мелкие частицы и микроорганизмы в отличие от макрофагов, способных к фагоцитозу более крупных частиц. 2. Осуществление нефагоцитарного типа бактерицидности путем секреции бактерицидных факторов, в т.ч. ферментов, бактериостатических и бактериоцидных белков, активных метаболитов кислорода. 3. Поддержание тканевого гомеостаза. Основной формой существования этих клеток является тканевой пейтрофил. После миграции в ткани, в основном в РВНСТ, нейтрофилы регулируют функции других клеток и межклеточного вещества путем секреции медиаторов и ферментов (см. ниже регуляторную функцию). 3. Разрушение раковых клеток — участие в противоопухолевой защите организма. 4. Секреторная и регуляторная функции — выделение медиаторов, регулирующих другие клетки, состояние межклеточного вещества, иммунные реакции, репаративиые процессы и др. Нарушения функций нейтрофилов могут проявляться в нарушениях хемотаксиса, угнетении фагоцитарной активности и нефагоцитарной бак-терицидности, в том числе и способности к респираторному взрыву. Характеризуются рецидивирующими бактериальными и 1рибковыми инфекциями. Эозинофильные лейкоциты, или эозинофилы. Имеют округлую форму и диаметр 10—12 мкм (рис. 9.1). В мазке имеют размеры 12—17 мкм, т.е. несколько крупнее нейтрофилов. В периферической крови их содержится небольшое количество — 2—5%. Основная форма их существования -тканевой эозинофил, количество которых в 200—300 раз превышает число этих клеток в крови. В крови эозинофилы находятся только несколько часов (3 — 12). Продолжительность жизни их в тканях составляет около 10 дней. Ядро эозинофилов имеет обычно два сегмента, из-за большего количества эухроматина светлее, чем ядро нейтрофила. В периферической крови кроме сегментоядерных могут изредка встречаться палочкоядерные и юные эозинофилы. Характерный признак — наличие двух типов гранул. 1. Ацидофильные гранулы. Составляют 95% всех гранул эозинофилов. При электронной микроскопии эозинофильные гранулы имеют овальную форму, слоистое строение, часто кристаллоидную структуру (рис. 9.8). Содержат фермент пероксидазу и другие ферменты лизосом (т.е. это лизосо-мы). Гранулы также содержат: 1) главный основной белок эозинофилов (Major protein basic, MPB). Обладает сильным антигельминтным, анти-протозойным и антимикробным свойством. Токсичен для тканевых клеток. Вызывает сокращение гладкомышечных клеток и дегрануляцию тканевых базофилов, базофильных лейкоцитов, тромбоцитов; 2) эозинофильный ка-тионный белок; 3) эозинофильный нейротоксин. Действие его схоже с действием МРВ. Нейротоксин способен повреждать нервные клетки; 4) гиста-миназу — фермент, нейтрализующий гистамин и оказывающий антиаллергическое действие.  2. Азурофильные (неспецифические гранулы) содержатся в небольшом количестве (5%). Являются лизосомами и содержат кислую фосфата-зу и ряд других ферментов, характерных для лизосом. Кроме гранул, в цитоплазме эозинофилов содержатся органеллы общего значения, в том числе и элементы развитого цитоскелста. Благодаря последним клетки способны активно мигрировать. Имеются также трофические включения гликогена, жиров, многочисленные пузырьки. Эозинофилы способны к самостоятельному движению и фагоцитозу, однако их фагоцитарная активность в отношении бактерий ниже, чем у нейтрофилов. Хемотаксические воздействия на эозинофилы оказывают комплекс антиген—антитело, гистамин. Активированные эозинофилы дегранулируют. Содержимое гранул токсично для простейших и гельминтов, опухолевых клеток, а также здоровых клеток и тканей. Функции. 1. Участие в аллергических реакциях: захват комплекса антиген—антитело, а также медиаторов аллергических реакций и их разру-дегрануляции эозинофилов могут играть патогенную роль: так, при бронхиальной астме они вызывают сокращение гладких миоцитои бронхов малого калибра, а также действуют повреждакнце на клетки бронхиального и альвеолярного эпителия. Базофильноые лейкоциты, или базофилы. Имеют размеры 8—10 мкм. Количество их в крови 0,5—1%, т.е. это самая малочисленная разновидность гранулоцитов (см. рис. 9.1). В периферической крови базофилы циркулируют до 1 суток, а затем перемещаются в ткани. Строение и функции базофильиых лейкоцитов во многом схожи со строением и функциями тканевых базофилов (тучных клеток) РВНСТ (см. ниже). Их размеры примерно равны размерам нейтрофилов (10— 12 мкм). Клетки имеют слабодольчатое плотное ядро, которое, однако, содержат больше эухрома-тина, чем нейтрофилы и эозинофилы. В цитоплазме содержатся орга-неллы общего значения, элементы цитоскслета, отдельные пузырьки и гранулы двух типов (рис. 9.9). 1. Базофильные, специфические гранулы. Эти гранулы окрашиваются метахроматически, то есть не в тот цвет, который имеет краситель в растворе. Метахромазия гранул обусловлена гепарином и хондроитин-сульфатом. Гранулы могут маскировать ядра клеток. Кроме гепарина, в гранулах содержитсягистамин (а у грызунов — и серотонин), ферменты (протеазы, пероксидаза и др.). Размеры гранул до 1—2 мкм. 2. Азурофильные гранулы немногочисленны, являются лизосомами. При действии стресс-факторов происходит дегрануляция базофилов. При этом гепарин препятствует свертыванию крови, серотонин и гистамин повышают проницаемость капилляров, стимулируют сокращение гладких миоцитов. Функции. 1. Участие в аллергических реакциях — инактивация комплекса антиген—антитело. 2. Регуляция проницаемости капилляров. Гистамин может вызвать отек ткани, сдавливание сосудов, это снижает концентрацию антигена, препятствует распространению антигена в организме. 3. Фагоцитоз бактерий и других антигенов. 4. Стимулируя сокращение гладких миоцитов полых органов, например, кишечника, гистамин базофилов способствует механическому удалению из организма паразитов. При некоторых заболеваниях (кожная базофильная гиперчувствительность и др.) количество базофилов в тканях может резко увеличиваться, что сопровождается их усиленной дегрануляцией. Клинически это проявляется крапивницей с сильным кожным зудом, аллергическим ринитом, приступами бронхиальной астмы, а в тяжелых случаях — анафилактическим шоком. НЕЗЕРНИСТЫЕ ЛЕЙКОЦИТЫ Моноциты. Это самые крупные клетки крови. В крови их 6—8% от всех лейкоцитов (абсолютное число равно 2х10!) клеток), тогда как в тканях их число в 20—30 раз превышает содержание данных клеток в кровяном русле. 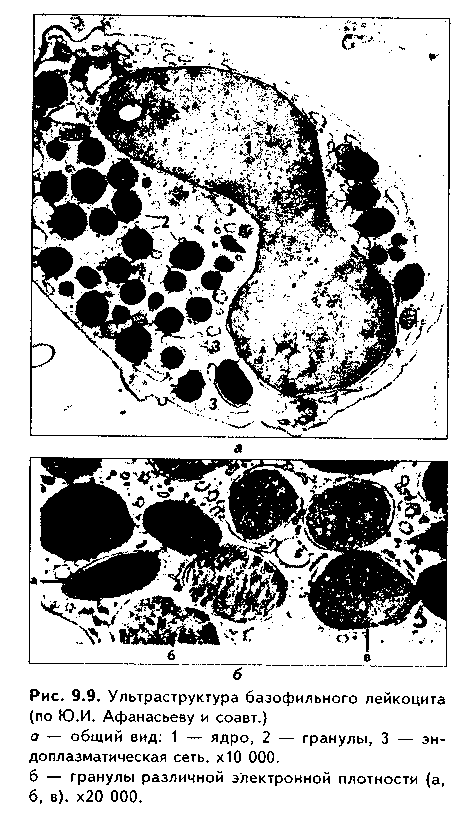 Моноциты имеют характерное строение. Их размеры в мазке составляют около 20 мкм (рис. 9.1). Цитоплазма слабобазофильна. В цитоплазме моноцитов могут быть мелкие азурофильные гранулы — лизосомы, содержится много вакуолей. Развиты гранулярная и агранулярная ЭПС, свободные рибосомы и полисомы, митохондрии, лизосомы, комплекс Гольджи (рис. 9.10). Хорошо развит цитоскелет, что обеспечивает подвижность клеток. Имеются включения гликогена. Азурофильные гранулы моноцитов дают реакцию на кислую фосфатазу и пероксидазу. Клетки имеют крупное, эксцентрично лежащее бобовидное, иногда дольчатое светлое ядро с небольшими ядрышками и пылевидным гетерохроматином. Из крови моноциты проникают в ткани, где могут некоторое время находиться в неизмененном состоянии, но обычно достаточно быстро превращаются в макрофаги. Это превращение заключается в увеличении размеров клеток, накоплении в них лизосом, других органелл, особенно белкового синтеза, изменении рецепторного аппарата и др. Макрофаги способны к вы-раженному фагоцитозу. Сами моноциты также могут фагоцитировать, но в меньшей степени, чем макрофаги. Кроме способности к фагоцитозу моноциты содержат антимикробные системы, позволяющие осуществлять нефагоцитарный тип бактерицидное -ти. К этим системам относят лизоцим, лактоферрин, катионные белки, миелопероксидазу, а также активные формы кислорода, появляющиеся при респираторном взрыве. Токсическим (и регуляторпым, вазоактивным) эффектом обладает также оксид азота (NO), вырабатываемый клетками 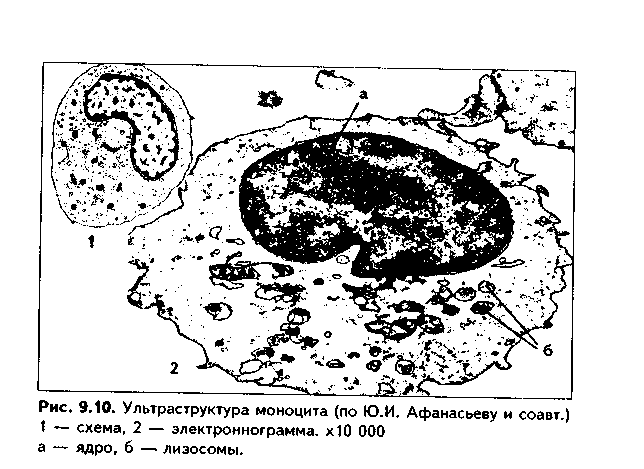 Моноциты относятся к системе мононуклеарных фагоцитов (СМФ). Кроме них в эту систему входят производные моноцитов — макрофаги разной локализации: гистиоциты рыхлой волокнистой соединительной ткани, клетки Купфера печени, остеокласты костной ткани, макрофаги селезенки, красного костного мозга, лимфоузлов, микроглия нервной ткани, клетки Лангерганса эпидермиса, альвеолярные, перитонеальные макрофаги. Клетки СМФ участвуют в фагоцитозе и иммунных реакциях, захватывают антигены, перерабатывают их (процессинг) и передают в высокоиммунной форме лимфоцитам (презентация), выделяют медиаторы, стимулирующие иммунные реакции, самостоятельно осуществляют разрушение чужеродных и опухолевых клеток (киллинг). Функции моноцитов. 1. Участие в неспецифических защитных реакциях путем фагоцитоза. 2. Участие в специфических (иммунных) защитных реакциях: процессинг и презентация антигенов лимфоцитам, выработка медиаторов иммунных реакций, киллинг (разрушение) чужеродных клеток. 3. Участие в противоопухолевой защите. 4. Регуляторная функция — синтез медиаторов (монокинов). 5. Участие в поддержании тканевого гомеостаза. Лимфоциты. Являются основными клетками иммунной системы. Их количество в крови равно 20—35%, т.е. это вторая по объему клеточная популяция периферической крови. Она представлена морфологически очень сходными, но функционально сильно различающимися клетками (рис. 9.1). Различия заключаются также в рецепторном репертуаре лимфоцитов. В крови лимфоциты находятся ограниченное время, после чего проникают в различные ткани и прежде всего заселяют ретикулярную ткань лимфоид-ных органов (селезенки, лимфоузлов, миндалин, аппендикса и др.), в которых являются основной клеточной популяцией. Из тканей лимфоциты вновь способны возвращаться в кровь. Этот процесс называется рециркуляцией лимфоцитов. Продолжительность жизни лимфоцитов резко варьирует и может составлять от нескольких часов до нескольких лет, причем в крови преобладают долгоживущие лимфоциты (70-75%).  По величине различают малые, средние и большие лимфоциты. Размеры их соответственно равны около 6, 8, 10 мкм (рис. 9.1). В крови преобладают малые лимфоциты (около 90%). Они имеют плотное округлое или бобовидное базофильное ядро и узкий ободок базофильной цитоплазмы. В цитоплазме содержится небольшое количество органелл общего назначения (|uic. 9.11): рибосом и полирибосом, митохондрий, встречаются элементы гладкой и гранулярной ЭПС, центриоли. Малые лимфоциты являя ются дифференцированными клетками, закончившими развитие в централь ных органах иммуногенеза (тимусе и красном костном мозге). Иногда такие лимфоциты определяют как "наивные" или "девственные" лимфоциты Они способны участвовать в иммунных реакциях организма только в результате специфических преобразовании первичного контакта с антигеном Эти преобразования сводятся к реакции бласттрансформации и последующей специфической дифференцировке (следовательно, лимфоциты как бы дважды проходят дифференцировку). Реакция бласттрансформации лимфоцитов (РБТЛ) — это превращение малых лимфоцитов в бластные клетки. РБТЛ включает в себя ряд морфологических и биохимических превращений лимфоцитов. В клетках резко повышается синтез ДНК (что регистрируется усилением включения меченого 3И-тимидина), увеличиваются размеры и количество ядрышек, размеры ядра, в котором начинает преобладать эухроматин. Одновременно цитоплазма клеток становится резко базофилыюй, увеличивается в объеме. В цитоплазме нарастает количество органелл: свободных рибосом, гранулярной ЭПС, объем комплекса Гольджи и лизосомального аппарата также возрастает. Изменяется рецепторный аппарат, в том числе репертуар CD-молекул цитолеммы (дифференцировочных молекул, англ. cluster differentiation). РБТЛ возникает, как указывалось, при контакте малых лимфоцитов со специфическим антигеном, а также действии на них митогенов и интерлейки-нов. Бласттрансформация лимфоцитов была впервые описана русским гистологом А.А. Максимовым в начале XX века. Она часто используется в диагностических целях для оценки иммунного статуса больных. Средние лимфоциты встречаются в крови в 10% случаев. Похожи на малые лимфоциты, от которых отличаются несколько большими размерами и более светлым ядром и большим объемом цитоплазмы. Большие лимфоциты в подавляющем большинстве являются бласт-ными клетками и в норме в периферической крови не встречаются (исключение — NK-клетки, см. ниже), локализуясь в органах иммунитета в зонах пролиферации. Характеризуются крупными размерами (до 18 мкм), светлым, с преобладанием эухроматина ядром, в котором видны крупные ядрышки, базофильной цитоплазмой с хорошо развитыми органеллами. Выделяют Т-лимфоциты (70% всех лимфоцитов крови), В-лимфоциты (10—20%) и "нулевые" лимфоциты (до 10%). Разделение лимфоцитов на Т-и В-популяции было предложено А. Ройтом (1969), название лимфоцитов основано на первых буквах органов, где они образуются. Т-лимфоциты дифференцируются из стволовой клетки в тимусе под влиянием его гормонов. Т-лимфоциты по функции делятся на Т-киллеры, Т-хелперы/индукторы, Т-супрессоры/цитотоксические и Т-лимфоциты памяти. Т-киллеры, или Т-эф-фекторы, участвуют в реакциях клеточного иммунитета (в последнее время эту субпопуляцию как изначально существующую не выделяют). Они рас-познают чужеродные клетки (клетки трансплантата, раковые клетки, клетки, зараженные внутриклеточными паразитами и вирусами и т.д.), прикрепляются к ним или же на расстоянии выделяют белки перфорины, которые повреждают мембрану этих клеток и вызывают их гибель. По современным данным, эти клетки могут также запускать в клетках-мишенях программу апоптоза. Т-хелперы/индукторы стимулируют реакции клеточного и гуморального иммунитета, Т-супрессоры/цитотоксические их подавляют. В последнее время считается, что эти клетки при активации их антигеном превращаются в Т-киллеры, т.е. Т-киллеры образуются только под индуктивным влиянием антигена. Т-лимфоциты памяти являются долгоживущими лимфоцитами, сохраняющими информацию об антигене и реализующими иммунные реакции при повторном контакте с ним. В-лимфоциты у птиц развиваются в бурсе Фабрициуса, а у человека и других млекопитающих — в красном костном мозге. В периферических органах иммуногенеза или в РВНСТ они после бласттрансформации превращаются в плазмоциты, которые вырабатывают антитела, инактивирую-щие антигены (т.е. участвуют в гуморальном иммунитете). В настоящее время установлено, что В-лимфоциты обладают способностью к процесси-рованию антигенов и презентации другим лимфоцитам. Существуют В-лимфоциты памяти, участвующие в иммунном ответе на повторное внедрение антигенов. Некоторые исследователи выделяют В-супрессоры, подавляющие иммунные реакции. Существует еще одна разновидность лимфоцитов, которая называется натуральными киллерами (NK-клетки). Они содержат в цитоплазме секреторные гранулы и часто называются большими гранулярными лимфоцитами (БГЛ). Количество этих клеток в крови составляет около 10% от всех лимфоцитов крови. Источник развития их неизвестен (возможно, это субпопуляция Т-лимфоцитов). Их основной, но не единственной функцией является участие в противоопухолевом иммунитете. "Нулевые" лимфоциты (0-лимфоциты) не несут маркеров ни T-, ни В-лимфоцитов. Это, очевидно, разнородная группа клеток, среди которых преобладают NK-клетки. Подробнее о функциях лимфоцитов, в том числе и NK-клеток, — см. "Основы частной гистологии". ЛЕЙКОЦИТАРНАЯ ФОРМУЛА. ГЕМОГРАММА Строение и состав периферической крови характеризуются достаточно жестким постоянством, красноречиво характеризуя гомеостаз организма. При этом в клинике наиболее часто используются такие показатели, как лейкоцитарная формула и гемограмма. Лейкоцитарная формула — этопроцентное содержание всех видов лейкоцитов периферической крови. Он;; выглядит так:  Примечание: цифрами дано процентное содержание лейкоцитов. Ю — юные нейт-рофилы (метамиелоциты); П — палочкоядерные, С — сегментоядерные нейтрофилы. Диагностическое значение лейкоцитарной формулы велико. Например, в клинике существуют такие понятия, как сдвиг лейкоцитарной формулы влево и вправо. Сдвиг влево — появление большого числа юных и палочко-ядерных (в первую очередь нейтрофилы шх) лейкоцитов. Наблюдается при воспалении, когда из красного костного мозга для реализации воспалительной реакции экстренно выбрасываются недостаточно зрелые формы лейкоцитов. Сдвиг вправо — отсутствие молодых форм нейтрофилов. Имеет место при нарушении нейтрофилопоэза. При лейкозах отмечается так называемый "лейке-мический провал" (hiatus leukemicus), когда одновременно возрастает количество незрелых и зрелых форм лейкоцитов при отсутствии переходных форм. Увеличение эозинофилов (эозинофилия) отмечается при аллергических реакциях, глистных инвазиях и других паразитарных заболеваниях. Снижение их числа имеет место при острых инфекциях, лечении глюкокорти-коидами и адренокортикотропином. Количество базофилов может быть увеличено (базофилия) при кожной базофилыюй гиперчувствительности, бронхи&тьной астме, а уменьшается при воспалительных процессах, после облучения, тиреотоксикозе и ряде заболеваний крови. Гемограмма — это абсолютное содержание форменных элементов крови, кроме этого, в гемограмму входят такие показатели: содержание ретикулоцитов; скорость оседания эритроцитов (СОЭ); содержание гемоглобина; гематокрит; а также лейкоцитарная формула. Ниже приводятся данные гемограммы без лейкоцитарной формулы (ее данные см. выше) 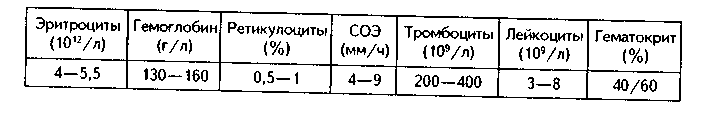 ВОЗРАСТНЫЕ ИЗМЕНЕНИЯ КРОВИ В постнатальном онтогенезе существенно изменяются практически все морфологические показатели крови. Врач любой специальности должен знать возрастные особенности строения крови. Эритроциты. Количество у новорожденных увеличено до 6—7х10,2/л, к 2-недельному возрасту достигает уровня взрослых и продолжает снижаться до минимума к 3—6-му месяцам жизни (физиологическая анемия). Дефинитивного количества их содержание достигает к половому созреванию. У новорожденных отмечаются анизоцитоз и ретикулоцитоз (увеличение количества ретикулоцитов). При старении количество эритроцитов может снижаться. Лейкоциты. При рождении отмечается физиологический лейкоцитоз (до 10—ЗОхЮУл). Дефинитивный уровень устанавливается к 14-ти годам. Имеют место физиологические перекресты, обусловленные изменениями содержания нейтрофилов и лимфоцитов. У новорожденного процентное содержание этих форм лейкоцитов примерно равно их уровням у взрослого. Первый перекрест отмечается на 3—4-е сутки жизни. К этому времени содержание клеток из-за падения доли нейтрофилов и повышения лимфоцитов уравнивается. Дальнейшие изменения ведут к тому, что к 1—2 годам жизни содержание нейтрофилов равно 25%, а лимфоцитов — 65%. В последующие 2—3 года наблюдается обратный процесс, и в 4 года наблюдается второй перекрест. К 14-ти годам показатели соответствуют таковым у взрослых людей. При старении могут наблюдаться снижение как абсолютного содержания лейкоцитов, так и сдвиги в лейкоцитарной формуле (отсутствие молодых форм нейтрофилов, снижение и отсутствие эозинофи-лов и др.) ЛИМФА Лимфа представляет собой продукт интерстициальной (внутритканевой) жидкости. Образуется путем фильтрации плазмы из кровеносных капилляров и венул, чему способствует высокое гидростатическое давление в интерстици-альном пространстве и различия в онкотическом давлении. Это обеспечивает поступление из плазмы крови в лимфу определенного количества белков, возвращаемых с лимфой обратно в кровь. Лимфа состоит из плазмы лимфы и форменных элементов (рис. 9.12). Плазма лимфы похожа по составу на плазму крови. Форменны*1 элементы составляют не более 1% объема лимфы, в процентном отношении это 95% лимфоцитов, 5% гранулоцитов, 1% моноцитов, Могут встречаться единичные эритроциты, благодаря чему, а также присутствию фибриноген;: и других факторов свертывания, лимфа коагулирует. Функции лимфы. 1. Транспортная, метаболическая и трофическая функции — транспорт липидов, всосавшихся в кишечнике, пластического и энергетического материала. 2. Перераспределение жидкости в организме 3. Участие в регуляции выработки антител, защитная функция. 4. Регуля-торная функция: является каналом передачи иммунной информации, фер ментов, гормонов и других регуляторных факторов. 5. Возвращение белка из ткани в кровь и поддержание онкотического давления крови. |