МЯДЕЛЕЦ ОБЩ.ГИСТ.. Литература для медицинских вузов все факультеты о. Д. Мяделец основы цитологии, эмбриологии и общей гистологии
 Скачать 4.75 Mb. Скачать 4.75 Mb.
|

|
4. Медиаторная классификация синапсов. По химическому типу медиатора есть синапсы холинергические, аминергические (адренергические, серотонинергические, дофаминергические); пуринергичес-кие, аминокислотные (медиаторами являются аминокислоты: ГАМК, глицин, глутамат, аспартат и т.д.), пептидергические (см. ме-диаторную классификацию нейроцитов). СТРОЕНИЕ СИНАПСОВ. Любой синапс состоит из трех частей: пре-синаптического полюса с пресинаптической мембраной, синапти-ческой щели и постсинаптического полюса с постсинаптической мембраной. 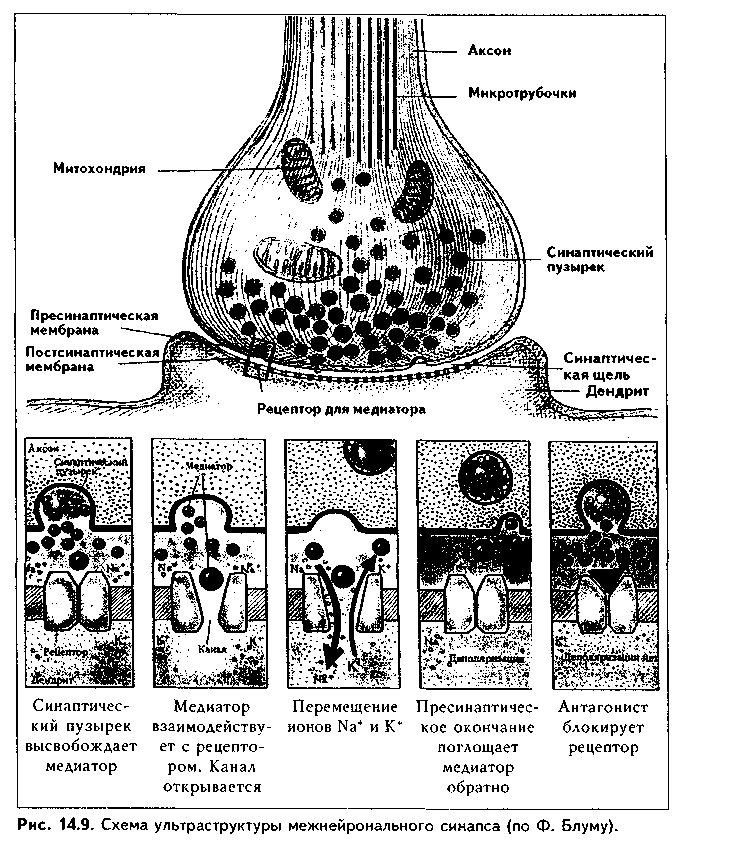 Электрические синапсы. Эти синапсы построены по типу нексусов: две мембраны (пре- и постсинаптическая) соседних нейронов тесно сближаются друг с другом до расстояния в 2 нм, и это место контакта пронизано многочисленными коннексонами. Следовательно, синаптическая щель в электрическом синапсе практически отстутствует. Коннексоны представляют собой своеобразную пору через обе мембраны, которая по краям ограничена особыми белковыми молекулами коннексинами. Коннексоны пропускают не только ионы щелочных металлов, играющих важную роль в формировании электрических потенциалов, но и молекулы с ММ 1000—2000. Поэтому кроме электрического сопряжения коннексоны дают возможность нейронам обмениваться метаболитами. В отличие от химических синапсов, в которых проведение сигнала несколько задерживается, в электрических синапсах импульс проводится практически без задержки и в обе стороны. Значение электрических синапсов неизвестно. Предполагают, что оно связано с необходимостью быстрого сопряжения нервных клеток. Химические синапсы. В отличие от электрических, химические синапсы передают нервные импульсы только в одном направлении и с задержкой (синаптическая задержка). Это наиболее распространенный у млекопитающих тин синапсов. Химические синапсы имеют все три отчетливо выраженные составные компоненты: нресинаптический и ностсинаптический полюсы и синаптичес-кую щель (рис 14.9) (в световом микроскопе синапсы видны в виде пугов-чатых утолщений на нейроцитах, см. рис. 13.1). В пресинаптическом полюсе находятся пресинаптические пузырьки с медиатором, митохондрии, агранулярная ЭПС, нейротрубочки и нейрофи-ламенты. Синаптические пузырьки имеют различное строение в зависимости от содержащегося в них медиатора. Так, пузырьки с ацетилхолином имеют мелкие размеры и электронно прозрачные. Синаптические пузырьки с но-радреналином крупнее и имеют в центре электронноплотную часть. Содержащие пептиды пузырьки имеют крупные размеры, плотную сердцевину и окружены периферическим светлым ободком. МОРФОЛОГИЧЕСКИЕ ОСНОВЫ БИОСИНТЕЗА И СЕКРЕЦИИ МЕДИАТОРА Медиаторные процессы в нейроне имеют несколько стадий: 1) синтез нейромедиатора; 2) его хранение (депонирование); 3) секреция нейромедиатора. Синтез нейромедиатора слагается из нескольких этапов. Вначале на гранулярной ЭПС в перикарионе синтезируются ферменты, осуществляющие биосинтез медиатора. Далее эти ферменты поступают в комплекс Гольджи, где "дозревают" и упаковываются в транспортные пузырьки. Эти транспортные пузырьки с помощью антероградного аксотока движутся в пресинаптический полюс. После поступления в пресинаптическую терминаль ферменты начинают синтезировать медиатор из предшественников, которые содержатся как в терминали, так и поступают из внеклеточного пространства. Далее медиатор упаковывается в пузырьки, мембраны для которых за счет механизма, похожего на механизм эндоцитоза, поставляет пресинаптическая мембрана. В пузырьках заключено около 10 000 молекул медиатора, что составляет квант. Вместе с медиатором в пузырьках всегда хранятся АТФ и некоторые катионы. Один нейрон может синтезировать несколько медиаторов. Например, существуют пептидхолинергические, пептидадренергические, пептидсеро-тонинергические и др. синапсы. Особенно часто встречаются нейроны, синтезирующие несколько видов медиаторов пептидной природы. В последнее время показано, что медиаторы, в первую очередь пептидные, могут синтезироваться и в перикарионе, а также по всему нейрону, откуда в транспортных пузырьках аксотоком доставляются в пресинаптический полюс. Депонирование медиатора осуществляется в пресинаптическом полюсе. Медиатор хранится в синаптических пузырьках. Секреция нейромедиатора осуществляется путем взаимодействия цитолеммы синаптических пузырьков и особых активных зон пресинаптической мембраны. Инициатором секреции является нервный импульс. В отсутствие последнего происходит секреция небольших доз медиатора, что вызывает в постсинаптической мембране спонтанные миниатюрные потенциалы. Их роль, очевидно, заключается в том, что при этом синапсы поддерживаются в состоянии постоянной готовности к ответу. На внутренней поверхности пресинаптической мембраны есть конусовидные плотные возвышения. Они соединяются друг с другом при помощи филамептов, поэтому вся внутренняя поверхность пресинаптической мембраны разделена на ячейки треугольной формы. Это зоны цитолеммы (активные зоны), через которые осуществляется секреция медиатора, выделяется содержимое синаптических пузырьков. Распространение нервного импульса по прссинаптическому полюсу ведет к открытию нотенциалзависимых кальциевых каналов, что увеличивает содержание кальция в пресинаптическом полюсе. Под действием кальция происходит взаимодействие актиновых и миози-иовых филамептов, а также запускается работа кинезинового механизма, что ведет к проталкиванию синаптических пузырьков в ячейки пресинаптической мембраны. Мембрана пузырьков сливается с мембраной пресинаптического полюса, и медиатор выделяется в щель, а затем идет к постсинаптической мембране, которая содержит рецепторы медиатора. Синаптическая щель имеет ширину около 30 нм. В ней содержатся особые элементы гликокаликса, которые обеспечивают адгезию пре- и постсинапти-ческого полюсов, а также целенаправленную диффузию медиатора. Некоторые авторы предполагают наличие в щели компонентов базальной мембраны. Постсинаптический полюс. Постсинаптическая мембрана имеет постсинаптическое утолщение за счет скопления под ней плотного филаментозного материала. В ней содержатся рецепторы медиатора. Взаимодействие медиатора с рецептором ведет к открытию ионных каналов в постсинаптической мембране, перераспределению ионов, деполяризации мембраны и возникновению нервного импульса. В тормозных синапсах, напротив, медиатор вызывает гиперполяризацию постсинаптической мембраны, что обеспечивает торможение. Медиаторами тормозных синапсов являются ГАМК и глицин. Кроме того, установлено, что и другие медиаторы (например, ацетилхолин, выполняющий возбуждающую функцию) могут вызывать тормозной эффект. Следовательно, медиатор может выполнять двойную функцию, а конечный эффект обусловлен характером рецепторов медиатора. После прекращения взаимодействия медиатора с рецептором он: 1) захватывается пресинаптической щелью и используется повторно (рециклинг медиатора); 2) поглощается окружающими глиальными клетками и разрушается ими; 3) расщепляется специальными ферментами (не все, а некоторые медиаторы, например, ацетилхолин, норадреналин). Обратные связи в синапсе. В последнее время установлено, что в синапсе существуют обратные связи, за счет которых обеспечивается постоянный контроль его работы. Обратные связи в синапсе осуществляются за счет нескольких механизмов. 1) "Пре-пре"-механизм. Осуществляется обратный захват медиатора из синаптической щели пресинаптической терминалью (рециклинг). При этом не только передается определенная информация из синаптической щели в пресинаптический полюс, но и происходит повторное использование медиатора. 2) "Пост-пост"-взаимодействие. Молекулы, выделившиеся из одного участка постсинаптического полюса, воздействуют на молекулы соседних участков этого же полюса. 3) "Пре-пост-пост-пре"-взаимодействие. Неспецифические продукты пре- и постсинаптического происхождения воздействуют как на пре-, так и на постсинаптическую мембраны. 4) "Пост-пре"-взаимодействие. Постсииаптические факторы оказывают действие на пресинаптическую мембрану. Обратные связи существуют как в нервно-мышечных, так и в нейро-нейрональных синапсах, обеспечивают четкую и ритмическую работу синапса, влияя на состояние как пре-, так и постсинаптического полюсов. Функции химических синапсов. 1) Передача возбуждения с одной нервной клетки на другую, обеспечение тем самым их связи в рефлекторных дугах; 2) Синапс обеспечивает поляризацию рефлекторных дуг, т.е. передачу нервного импульса в одном направлении; 3) Синапс является местом регуляции функций нервной системы; 4) Синапс — место, где обеспечивается и хранится нейрональная память; 5) Синапс играет важную роль в адаптивных перестройках нейрона. МЕХАНИЗМЫ АДАПТАЦИИ И КОМПЕНСАЦИИ НЕЙРОНОВ. В основе компенсаторно-приспособительных перестроек нейронов лежат механизмы внутриклеточной регенерации, в первую очередь, гипертрофия и гиперплазия органелл. При этом очень важная роль отводится процессам биосинтеза и секреции медиатора и перестройке работы синапсов. Можно выделить несколько основных позиций, определяющих протекание компенсаторно-приспособительных перестроек нейрона, связанных с синапсами. 1. Усиление выработки ферментов биосинтеза медиатора. 2. Усиление аксонного транспорта. 3. Усиление рециклинга медиатора. 4. Изменение активности ферментов деградации медиатора. 5. Изменение обратной связи в синапсе (усиление, ослабление). 6. Увеличение количества рецепторов на постсинаптической мембране. 7. Увеличение зоны контакта частей нейронов в синапсе. 8. Увеличение количества шипикового аппарата. 9. Увеличение количества функционирующих синапсов. ПОНЯТИЕ О РЕФЛЕКТОРНЫХ ДУГАХ Рефлекторная дуга — это цепь нейронов, связанных синапсами, обеспечивающая проведение импульса от рецептора к рабочему органу (мышце, железе). Различают простые и сложные рефлекторные дуги (рис. 14.10). Простые рефлекторные дуги состоят ил чувствительного и двигательного нейронов, связанных синапсом. Такие дуги состоят из следующих частей: рецептора, образованного дендритом чувствительного нейрона; дендрита, пери-кариона, аксона сенсорного нейрона; синапса сенсорного нейрона с эфферентным нейроном; дендрита, перикарио-на и аксона эфферентного нейрона; эффекторного (двигательного) нервного окончания. В сложных рефлекторных дугах большое количество нейронов, причем -их количество увеличивается за счет вставочных нейронов. Возбуждение по рефлекторной дуге передается только в одном направлении, поскольку'синапсы осуществляют их поляризацию. ОСНОВНЫЕ ПОЛОЖЕНИЯ НЕЙРОННОЙ ТЕОРИИ До конца XIX века существовала ретикулярная, или фибриллярная, теория организации нервной ткани, согласно которой она состоит не из клеток, а из истинного синцития. В 1891 году немецкий анатом В. Вальдейср выдвинул альтернативу этой теории и сформулировал нейронную теорию: нервная ткань состоит не из синцития, а из отдельных, дискретных нейронов. В разработке нейронной теории есть заслуга многих ученых-гистологов и анатомов. В частности, ряд интересных взглядов, не укладывающихся в теорию фибриллярного строения нервной ткани и противоречивших ей, был высказан в работах В. Гиса-старшсго и О. Фореля. Однако главная заслуга в создании нейронной теории принадлежит испанскому нейрогистологу, лауреату Нобелевской премии (1906) С. Рамону-и-Кахалу. Ему оппонировал другой знаменитый гистолог — итальянский ученый К. Гольджи. Несмотря на заблуждения К. Гольджи, отстаивавшего фибриллярную теорию, его вклад в развитие учения о нервной ткани был настолько велик, а противостояние сторонникам нейронной теории столь плодотворно, что совместно с С. Рамоном-и-Кахалем ему была присуждена Нобелевская премия. Оставался, однако, неясным вопрос механизмов коммуникации нейронов. Этот вопрос был разрешен работами английского физиолога У. Шерин-гтона, который ввел гипотетическое понятие "синапс" как место соединения двух клеток. Спустя четверть века теория синапса стала общепризнанной и окончательно утвердила нейронную теорию. В ее развитие большой вклад внесли также русские и советские гистологи: А.С. Догель (А.С. Догель не во всем принимал нейронную теорию, в частности, придерживался взглядов о фибриллярном строении сетчатки, но тем не менее его труды сыграли важную роль в утверждении нейронной теории), Б.И. Лаврентьев, А.А. Заварзин, Б.С. Дойников, Н.Г. Колосов, Г.И. Поляков и др. 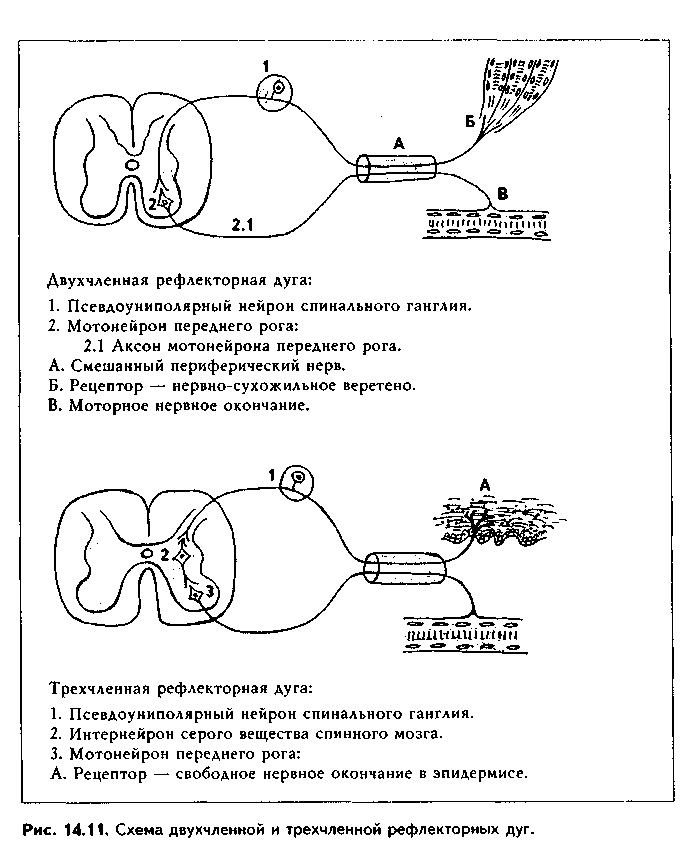 Основные положения нейронной теории были сформулированы в начале настоящего века. В выработке этих положений большая роль принадлежит С. Рамон-и-Кахалю, А.С. Догелю, Б.И. Лаврентьеву. Положения нейронной теории сводятся к следующему: 1. Структурно-функциональной, медиаторной и метаболической единицей нервной ткани и нервной системы является нейрон. 2. Нейрон — клетка, состоящая из перикариона, аксона, дендритов и их терминальных ветвлений. 3. Функционирование нейронов возможно только при тесной интеграции их с различными видами нейроглии. 4. Нейроны взаимодействуют друг с другом при помощи синапсов — специализированных межклеточных контактов. 5. Совокупность нейронов, связанных синансами, формируют рефлекторные дуги — основной субстрат нервной системы. 5. Возбуждение в синапсах и в рефлекторных дугах передается только в одном направлении.                |