МЯДЕЛЕЦ ОБЩ.ГИСТ.. Литература для медицинских вузов все факультеты о. Д. Мяделец основы цитологии, эмбриологии и общей гистологии
 Скачать 4.75 Mb. Скачать 4.75 Mb.
|

|
Глава 4 ЦИТОЛОГИЯ. СТРОЕНИЕ И ФУНКЦИИ ЯДРА. ЖИЗНЕННЫЙ ЦИКЛ КЛЕТКИ. РЕАКТИВНЫЕ ИЗМЕНЕНИЯ КЛЕТОК. СТАРЕНИЕ И СМЕРТЬ КЛЕТКИ СТРОЕНИЕ И ФУНКЦИИ КЛЕТОЧНОГО ЯДРА Ядро клетки является ее важнейшим структурным компонентом. Его функции следующие: 1. Хранение наследственной информации в молекулах ДНК хромосом. 2. Реализация наследственной информации путем контроля в клетке синтетических процессов, а также процессов воспроизводства и гибели (апоптоза). 3. Воспроизводство и передача генетической информации при делении клетки. 4. Контроль и регуляция структурно-функционального состояния цитоплазмы, клеточной оболочки, циторецепторов. Количество ядер, их форма, величина зависят от вида клетки и ее функционального состояния. Наиболее часто встречаются одноядерные клетки, однако у некоторых клеток (например, гепатоцитов и др.) в связи с интенсификацией функций может встречаться несколько ядер. Известны гистологические структуры (например, симпласты в поперечнополосатой мышечной ткани), для которых многоядерность является постоянным признаком. Форма ядер, как правило, зависит от формы клеток. Ядро может быть уплощенным в плоских, округлым в кубических, эллипсоидным в призматических клетках. Встречаются сегментированные, палочковидные, лопастные ядра. Расположение ядра также может быть различно: они могут лежать в центре клетки, эксцентрично, в базалыюй части. Размеры ядра в целом зависят от функционального состояния клетки: в функционально активных клетках ядро имеет крупные размеры и наоборот. Крупные размеры характерны также для полиплоидных ядер. В организме человека встречаются так называемые постклеточные структуры, иногда также неправильно называемые клетками: зритроциты, роговые чешуйки эпителия кожи, кровяные пластинки (тромбоциты). В них отсутствуют ядра, которые теряются в ходе специфической дифференцировки. В постклеточных структурах подавляющее большинство характерных для клетки процессов отсутствует, они в течение определенного времени выполняют одну или несколько функций, а затем гибнут. В интерфазной клетке ядро состоит из 4 компонентов (рис. 4.1): 1. Хроматин (как часть хромосом). 2. Ядрышко. 3. Кариолемма. 4. Кариоплазма. 1. ХРОМАТИН. Хроматином называется интерфазная форма существования хромосом. Структурное состояние хромосом существенно меняется в интерфазных ими-тотически делящихся клетках. В интерфазе хромосомы находятся в частично или полностью деконден-сированном состоянии. При этом большая их часть становится невидимой в световом микроскопе. Области деконденсации хромосом являются активными, здесь идет транскрипция ДНК. Такие области называются эухроматином. Конденсированный, или плотный хроматин имеет выраженную базофилию и виден в микроскопе. Эти неактивные участки хромосом иначе называются гетерохроматином. В  световом микроскопе гетерохроматин иидеи и форме гранул или глыбок, окрашенных основными красителями в характерный для красителя цвет и распределенных но ядру или относительно равномерно, или зонально. Иногда распределение гетерохроматина создает картину спин колеса (в плазмоцитах). Часть гетерохроматина прилежит к кариолемме — примем-бранный хроматин, — а также сосредоточена вокруг ядрышек — иеринукле-олярный хроматин. Гетерохроматин делится на два вида: 1. Конститутивный хроматин — это такой гетерохроматин, с которого никогда ни в одной клетке не идет считывание информации и виде и-РНК. В хромосомах это обычно области вблизи центромеров. 2. Факультативный гетерохроматин — это хроматин, количество которого заметно варьирует в разных клетках: его совсем мало в эмбриональных клетках, а по мере дифференцировки клеток содержание этого хроматина увеличивается. В синтезирующих белок клетках количество факультативного хроматина снижено. Поскольку красителями окрашивается только гетерохроматин, то степень окраски ядра зависит от его количества. Темноокрашенное ядро обычно характерно для функционально неактивной клетки. При активации клетки соотношение эухроматин/гетерохроматин изменяется в пользу эухроматина, и ядра функционально активных клеток светлые, слабоокрашенные. 2. ЯДРЫШКО. Это плотный структурный компонент ядра. В клетке может быть от одного до нескольких ядрышек. Ядрышко — это совокупность участков 10 хромосом (13, 14, 15, 21, 22 пары) (рис. 4.2, а). Эти участки называют ядрышковыми организаторами. Они находятся в области вторичных перетяжек хромосом и представлены многочисленными копиями генов рибосомальных РНК (рРНК). Следовательно, в ядрышках с ДНК ядрышковых организаторов происходит считывание информации в виде рибосомальной РНК. В световом микроскопе ядрышко определяется как плотноокрашенная основными красителями глобула размером от 1 до 3 мкм, не имеющая оболочки. Располагается как в центре ядра, так и эксцентрично. Интенсивно окрашивается на рибонуклеопротеиды. Размеры ядрышка тем больше, чем выше функциональная активность клетки. В электронном микроскопе ядрышко состоит из двух основных частей: фибриллярного (представлен первичными цепями рибосомальной РНК) и гранулярного (предшественники рибосом). Иногда выделяют третий, аморфный компонент ядрышка, который представляет собой собственно ядрыш-ковые организаторы. Ядрышко подвергается характерным изменениям в митотическом цикле (рис. 4.2, б). Во время митоза оно исчезает, потому что хромосомы спирализируются и расходятся, прекращается синтез РНК на ядрышковых организаторах. При этом ядрышко постепенно распадается на 10 частей (столько же, сколько и хромосом, его образующих), которые постепенно исчезают. После митоза ядрышко вновь восстанавливается: вначале образуется 10 мелких ядрышек; они сливаются и образуют одно—два ядрышка. 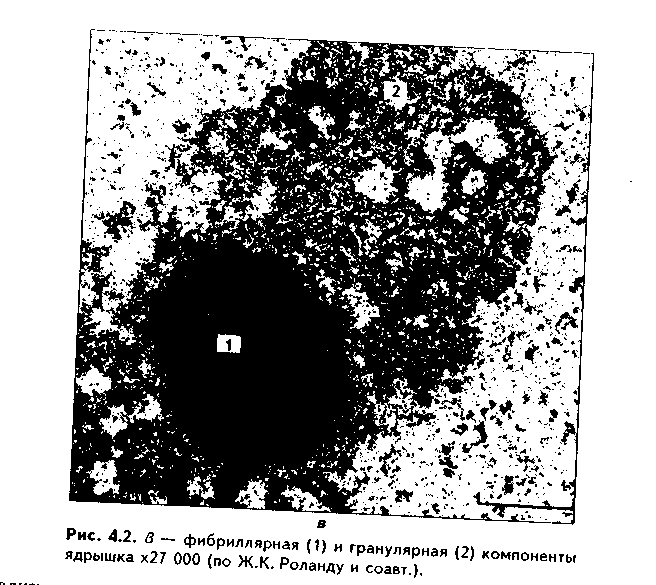 Функции ядрышка — синтез рибосомальной РНК и образование рибосом. При транскрипции генов ядрышковых организаторов вначале образуется гигантская молекула-предшественница рРНК. Она связывается с белками, синтезированными в цитоплазме и поступившими в ядро. Образуются рибопуклеопротеиды (РНП), которые подвергаются растеплению на более мелкие фрагменты, соединяющиеся с добавочными молекулами белка. Одна часть этих фрагментов превращается в большие, другая часть — в малые субъединины рибосом. 3. ЯДЕРНАЯ ОБОЛОЧКА, или кариолемма. На светомикроскопичес-ком уровне она видна как тонкая пластинка, окружающая ядро. В электронном микроскопе состоит из двух мембран, которые имеют такое же строение, как все биологические мембраны (рис. 4.3). Наружная мембрана переходит в мембраны эндоплазматической сети. На ней могут быть рибосомы. Со стороны цитоплазмы наружная мембрана окружена сетью промежуточных виментиновых филамеитов. Между двумя мембранами есть перинуклеарное пространство шириной 20—40 им. Оно является аналогом полостей гранулярной ЭГТС и может содержать продукты белкового синтеза.  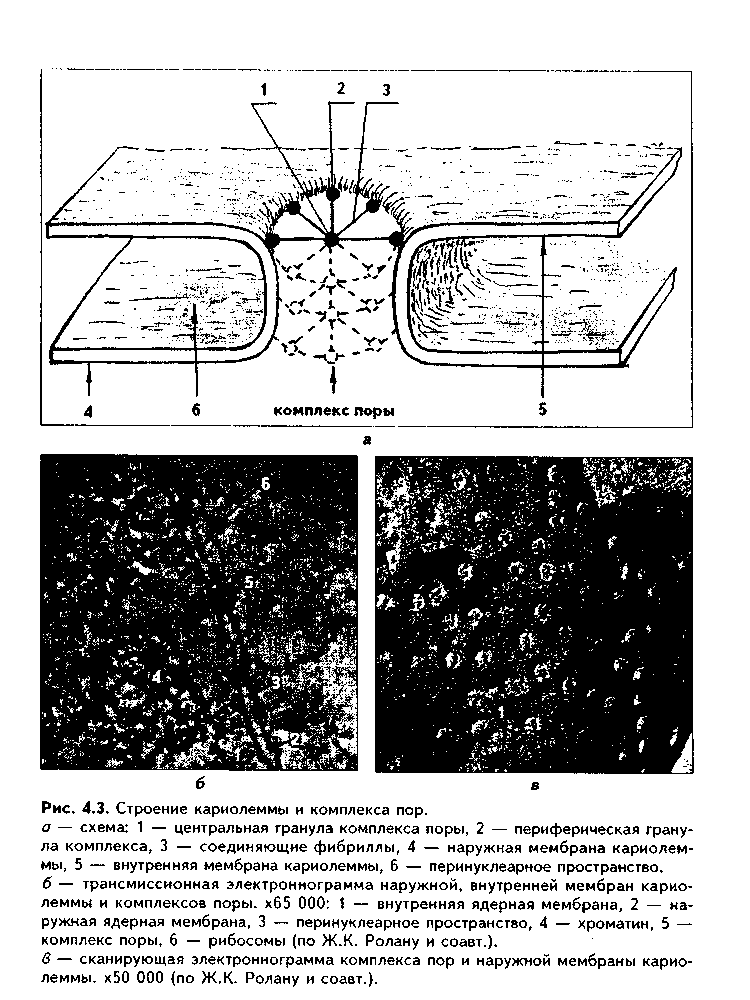 Внутренняя мембрана кариолеммы гладкая. При помощи стрруктурных белков она связана с плотно прилежащей к ней ламиной, или ядерной пластинкой, которая имеет толщину до 300 нм и состоит из сгущения промежуточных филаментов. С ламиной контактируют промежуточные фи-ламенты, формирующие в ядре фибриллярную сеть и образующие кариос-келет. Ламина поддерживает форму ядра, участвует в организации пор, способствует упорядоченному расположению хроматина. Она также участвует в формировании кариолеммы при делении клеток. Две ядерные мембраны в отдельных участках переходят одна в другую. Эти места являются порами кариолеммы (рис. 4.3). В порах находятся гранулярные и фибриллярные структуры, которые вместе образуют комплекс поры. По краю поры лежат 8 гранул, а в центре находится центральная гранула. К ней от периферических гранул идут фибриллы. Формируется структура, похожая на колесо со спицами. В комплексе поры имеется три таких структуры, которые лежат на разных уровнях, формируя три этажа. Гранулы пор связаны с белками ламины, участвующей в их организации. В комплексе поры содержатся особые рецепторы, распознающие поступающие в ядро белки и осуществляющие их активный перенос. Число пор зависит от метаболической активности клеток: чем выше синтетические процессы, тем выше содержание пор. В среднем в ядерной оболочке содержится 2000—4000 пор. В сперматозоидах ядерные поры полностью отсутствуют. Функции кариолеммы: 1. Разграничительная. 2. Защитная. 3. Регуляция транспорта веществ, в том числе и рибосом, из ядра в цитоплазму и наоборот. Комплекс пор играет в этом наибольшее значение (роль диафрагмы и активного транспортера). 4. ЯДЕРНЫЙ СОК - КАРИОПЛАЗМА. Это жидкий компонент ядра. Представляет собой коллоидный раствор сложных белков, углеводов, нуклеотидов. В состав кариоплазмы входят также различные ионы и метаболиты. Среди белков наибольшее значение имеют гистоны, ферменты, структурные белки. Функции кариоплазмы: 1. Создает микросреду для всех структур ядра, в которой может происходить быстрая диффузия метаболитов. 2. Перемещение рибосом, м-РНК и т-РНК к ядерным норам. ХРОМОСОМЫ. Хромосомы видны полностью только в митозе. Наиболее удобно изучать их в метафазе (метафазн^" хромосомные пластинки). Основными химическими элементами хромосимы являются ДНК и белки (рис. 4.4). Комплекс ДНК с белками (в основном с гистонами) формирует фибриллярную структуру — элементарную хромосомную фибриллу, имеющую нуклеосомную организацию. Каждая нуклеосома представляет собой комплекс из 8 молекул гистонов (гистоновый октамер). Вокруг него молекула ДНК образует около 2 оборотов. Участки ДНК, связывающие соседние нуклеосомы, называются линкерной ДНК. Следующим уровнем организации хромосомы является нуклеомерная организация, или уровень хроматиновой фибриллы. В ней нуклеосомы объединяются в нуклеомеры, причем каждый нуклеомер состоит из 8—10 нуклеосом и имеет диаметр около 30 нм. Хромосомы образованы хроматиновыми фибриллами (хрома-тидами) в интерфазе. В ходе последующей упаковки нуклеомер подвергается суперспирализации и превращается в хромомер, содержащий петельные домены. Петельный домен имеет диаметр до 300 им и соответствует одному или нескольким генам. Хромомер далее за счет сунерспирализации укорачивается, образуются конденсированные хромосомы, видимые только в митозе клетки. 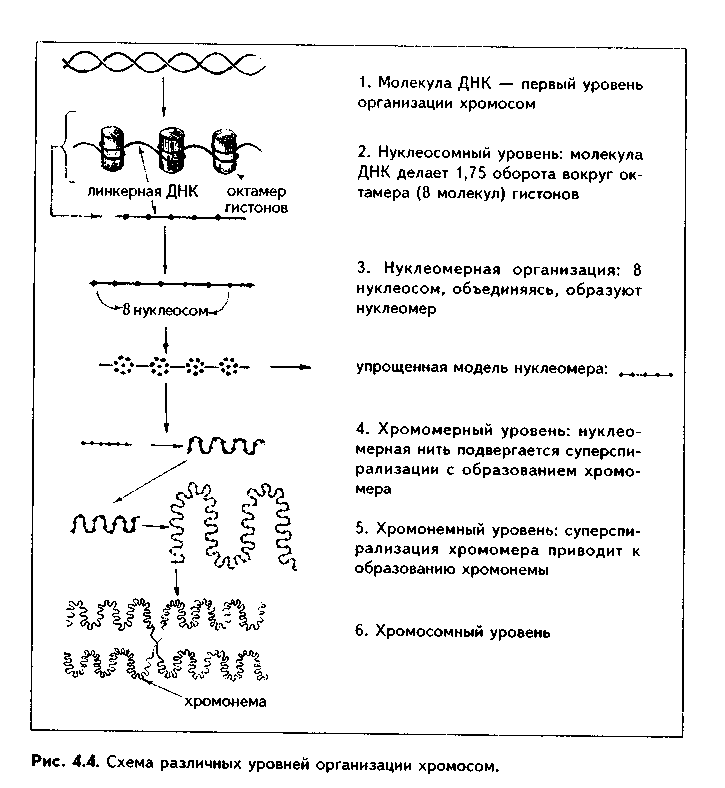 МОРФОЛОГИЯ И КЛАССИФИКАЦИЯ ХРОМОСОМ Каждая хромосома в световой микроскоп выглядит в виде палочки (рис. 4.5). У большинства хромосом можно увидеть первичную перетяжку — центромер, или кинетохор. Он делит хромосомы на два плеча. Если длина плеч одинакова, такие хромосомы называются метацентрическими. Если одно плечо больше, то хромосомы являются субметацентрическими. Хромосомы с очень коротким одним плечом называются акроцентрически-ми. Иногда на хромосомах есть вторичные перетяжки, отделяющие от хромосомы маленький участок — сателлит. В области вторичных перетяжек находятся ядрышковые организаторы. Согласно Денверской классификации хромосом (Денвер, США, 1960), учитывающей размеры хромосом, расположение первичных, вторичных перетяжек и наличие спутника, все хромосомы делятся на 7 групп (А, В, С, D, E, F, G) (рис. 4.5, б). Существует также Парижская (1971) классификация хромосом. В ее основу положена дифференциальная окраска хромосом некоторыми красителями. Эта окраска выявляет в хромосомах чередующиеся светлые и темные полосы (гетеро- и эухроматиновые районы), уникальные для каждой пары хромосом. Дифференциальная окраска позволяет достоверно отличить одну пару хромосом от другой. Все хромосомы вместе образуют кариотип. Среди хромосом есть соматические, или аутосомы (рис. 4.5, б), и половые хромосомы. Соматические хромосомы образуют гомологические (похожие) пары. У человека таких пар 22. Половые хромосомы различаются в мужском и женском организме. В женском организме есть две Х-хромосомы, в мужском — половые хромосомы различные, X и Y, причем Y-хромосомы определяют пол мужчины. СПОСОБЫ РЕПРОДУКЦИИ КЛЕТОК Универсальным механизмом размножения клеток является митоз, или непрямое деление. Разновидностями митоза являются мейоз и эндо-митоз. Иногда как самостоятельный способ репродукции рассматривают амитоз, или прямое деление. Однако в последнее время большинство цитологов отрицают существование амитоза у эукариотических клеток. МИТОЗ. Это непрямое деление клетки, связанное с изменениями ее ядра. В митозе имеется 4 фазы: профаза; метафаза; анафаза; телофаза (рис. 4.6, 4.7). В ПРОФАЗУ происходят следующие события: 1. В результате спирализации и конденсации хроматина становятся видны хромосомы. Каждая хромосома состоит из двух лежащих рядом сестринских хроматид. 2. Исчезает ядрышко, т.к. на ядрышковых организаторах прекращается синтез р-РНК и они расходятся в связи с конденсацией хромосом. 3. Из микротрубочек цитоплазмы формируется веретено деления. Центрами его организации становятся разошедшиеся к полюсам центриоли. Микротрубочки веретена деления прикрепляются к центромерам хромосом, в области которых из особых белков формируются кинетохоры. В дальнейшем кинето-хоры сами могут служить центрами организации микротрубочек. 4. Распадается на мелкие фрагменты, превращается в мембранные пузырьки и становится неотличимой от ЭПС ядерная оболочка. По-ровый комплекс и ламина распадаются на субъединицы. МЕТАФАЗА. Все хромосомы располагаются в экваторе клетки и удерживаются в этом положении микротрубочками веретена деления. Сестринские хроматиды отходят друг от друга, разделяясь щелью, но остаются соединенными в области центромеры. Хромосомы формируют метафазную пластинку, или материнскую звезду. АНАФАЗА. Сестринские хроматиды, из которых состоят хромо-сомы, отделяются друг от друга и в области центромеры и начинают расходиться к полюсам клетки со скоростью до 1 мкм/мин. Анафаза обычно длится несколько минут.   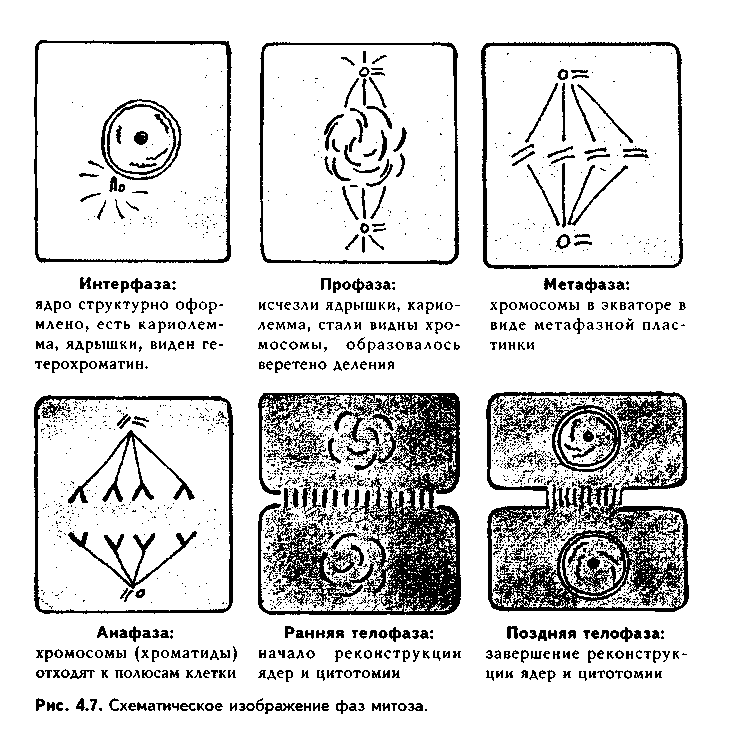 Механизм движения хроматид к полюсам не совсем ясен. Предполагают, что сигналом к движению является резкое повышение в гиалоплазме концентрации ионов кальция. Возможно, причина движения заключается в деполимеризации микротрубочек веретена с конца, прикрепленного к кинетохорам. По другим представлениям, оиа кроется во взаимодействии таких сократимых белков, как актин, миозин и динеин, которые сосредоточиваются вокруг веретена деления. Разошедшиеся к полюсам сестринские хроматиды формируют дочерние звезды. ТЕЛОФАЗА. Когда разделенные дочерние хроматиды подходят к полюсам, кинетохорные трубочки исчезают. Вокруг каждой группы дочерних хроматид из мембранных пузырьков и агранулярной ЭПС образуется новая ядерная оболочка, а из имеющихся в цитоплазме субъединиц — норовые комплексы и ламипа. Конденсированный хроматин начинает деспирализо-ваться, разрыхляться. Появляются ядрышки. Происходит распределение органелл между клетками. Затем, благодаря актииовым филаментам, в центре клетки по периметру образуется сократимое кольцо (рис. 4.7, 4.8). Оно постепенно сжимается и образует борозду деления, которая углубляется и в конце концов разделяет материнскую клетку па две клетки. Это явление называется цитотомией. 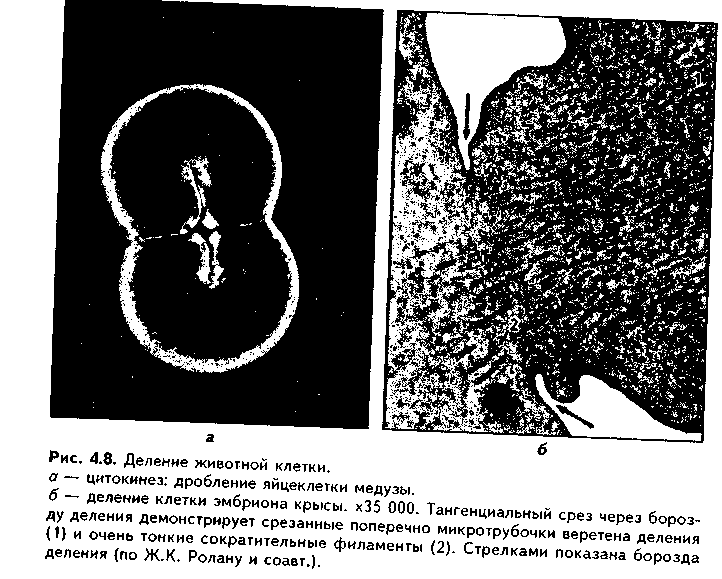 Наряду с описанным нормальным митозом могут наблюдаться атипичные и патологические митозы. При них может иметь место неравномерное распределение генетического материала между дочерними клетками — анэуплоидия. Могут наблюдаться также аномалии хромосом — хромосомные аберрации, часто возникающие после рентгеновского облучения. Патологические митозы характерны для опухолевых клеток. ЭНДОМИТОЗ — это вариант митоза, когда редупликация хромосом не заканчивается образованием двух клеток. Есть несколько вариантов эн-домитоза, отражающих степень "продвинутое™" митоза: 1. ПОЛИТЕНИЯ — явление, при котором в результате редупликации ДНК происходит увеличение размеров хромосом во много раз. Имеет место у беспозвоночных животных. 2. ПОЛИПЛОИДИЯ — увеличение количества хромосом, обычно кратное двум. В полиплоидных клетках в последующем может происходить разделение (сегрегация) геномов, и такие клетки распадаются на несколько клеток с диплоидным набором хромосом. Эти изменения некоторые исследователи рассматривают как проявление амитоза. 3. Образование ДВУЯДЕРНЫХ И МНОГОЯДЕРНЫХ КЛЕТОК. Они возникают тогда, когда ядро делится, но цитотомия не происходит. Многоядерные клетки в последующем могут путем цитотомии разделиться с образованием одноядерных (вариант амитоза). Эидомитоз в конечном счете приводит к увеличению размеров клетки и ее функциональных возможностей, поэтому его можно рассматривать как механизм приспособления или адаптации клетки к изменяющимся условиям внешней среды. АМИТОЗ, или прямое деление клетки. В последнее время большинство исследователей отрицают его существование и значение для организма. Поэтому во многих руководствах амитоз вообще не рассматривается как способ репродукции клеток. Амитоз — деление клетки без изменений со стороны хромосомного аппарата. Он проходит путем простой перетяжки ядра и цитоплазмы без выявления хромосом и образования веретена деления. Одной из форм амитоза является сегрегация геномов — множественная перешнуровка полиплоидного ядра с образованием мелких дочерних ядер с обычным генотипом. Те исследователи, которые допускают существование амитоза, различают реактивный амитоз (ответная реакция на внешние факторы), патологический амитоз (в условиях патологии), регенераторный амитоз (при регенерации поперечно-полосатой мышечной ткани), дегенеративный амитоз (в стареющих клетках). МЕЙОЗ. Это деление половых клеток, вариант митоза. При помощи мейоза образуются клетки с гаплоидным набором хромосом. Мейоз состоит из двух последовательных митотических делений: МЕЙОЗ I и МЕЙОЗ И. Мейоз I называют редукционным делением, т.к. в нем происходит редукция, уменьшение хромосомного набора в два раза. Мейоз I имеет сложную профазу, состоящую из 5 периодов, или фаз: — ЛЕПТОТЕНА — хромосомы приобретают вид длинных тонких нитей; — ЗИГОТЕНА — в ней происходит конъюгация гомологичных хромосом; — ПАХИТЕНА — в эту фазу хромосомы укорачиваются и утолщаются; — ДИПЛОНЕМА — характеризуется расщеплением хромосом на две половинки — хроматиды. Образуются тетрады, состоящие из четырех хроматид; — ДИАКИНЕЗ — хромосомы сильно укорачиваются в результате спи рализации и отходят друг от друга. Дальнейшие фазы мейоза I (метафаза, анафаза, телофаза) такие же. как в митозе, но к полюсам отходят не хроматиды, а целые хромосомы. Это и приводит к редукции хромосомного набора. В мейозе II к полюсам отходят хроматиды, как в митозе. Подробнее о мейозе — в разделе "Эмбриология", см. рис. 5.5. ЯДЕРНО-ЦИТОПЛАЗМАТИЧЕСКОЕ ОТНОШЕНИЕ КАК ПОКАЗАТЕЛЬ ФУНКЦИОНАЛЬНОГО СОСТОЯНИЯ КЛЕТКИ Отношение площади или объема ядра к площади или объему цитоплазмы называется ядерно-цитоплазматическим отношением (ЯЦО). Ядер-но-цитоплазматическое отношение показывает, в каком состоянии находится клетка. Если это отношение равно или больше 1, это значит, что в клетке большое ядро и мало цитоплазмы. Такое отношение могут иметь стволовые клетки, малые лимфоциты, стареющие клетки. Эти клетки функционально неактивны, однако обладают способностью делиться, например, стволовые клетки. Наоборот, клетки, у которых ЯЦО меньше 1, имеют большой объем цитоплазмы и, следовательно, большое количество органелл. Они высокодифференцированы и способны активно функционировать. МИТОТИЧЕСКИЙ ЦИКЛ. ЖИЗНЕННЫЙ ЦИКЛ КЛЕТКИ Митотическнй цикл — это время от одного до второго деления клетки. Его подразделяют на собственно митоз и интерфазу. В свою очередь, интерфаза делится на 3 периода (рис. 4.9, 1): 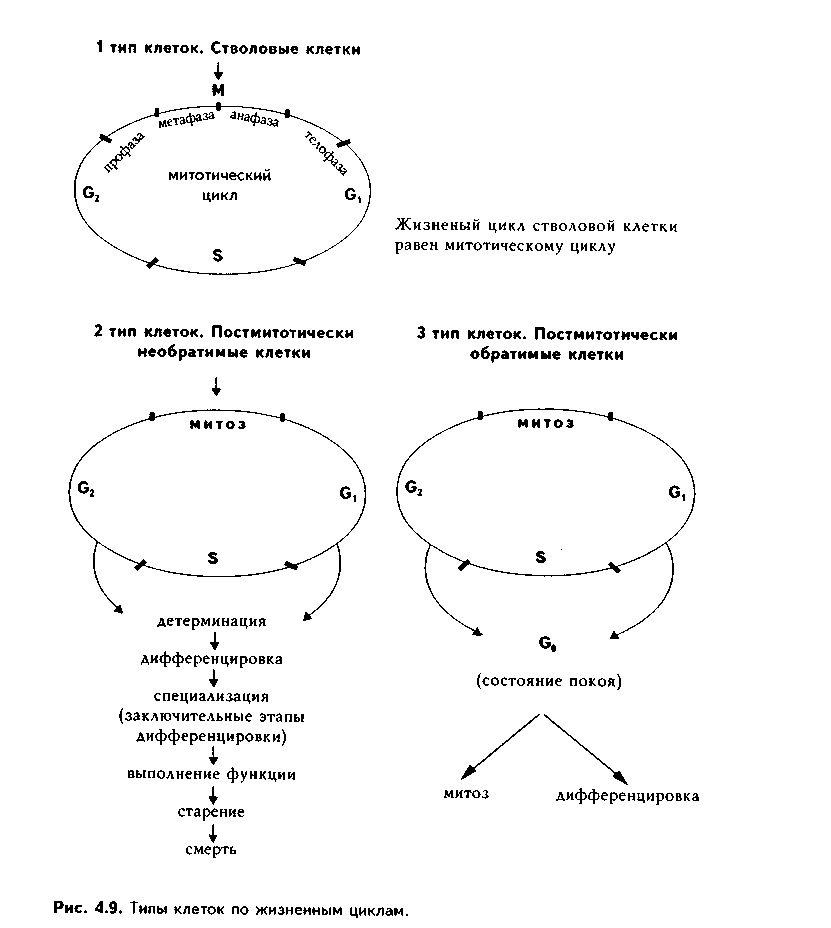 1. G,-период. В нем активируются обменные процессы, необходимые для синтеза ДНК. Характеризуется ростом клеток, синтезом белка и РНК. Клетка восстанавливает нужный объем органелл и достигает обычных размеров. Синтезируются также специальные белки-активаторы S-периода. 2. S-период — период синтеза, удвоения ДНК в ядре, хромосомы полностью реплицируются. Одновременно удваиваются центриоли. 3. С2-период — синтез и-РНК, р-РНК, белков тубулинов, из которых синтезируется веретено деления. Полностью созревают дочерние центриоли. Запасается энергия. Затем наступает М-период, или собственно митоз. ЖИЗНЕННЫЙ ЦИКЛ — это время от одного деления до второго или до смерти клетки. Есть три основных вида тканевых клеток, различающихся по жизненному циклу (рис. 4.9): 1. Стволовые клетки. Эти клетки способны к постоянному делению митозом. За счет них поддерживается тканевой гомеостаз. Жизненный цикл таких клеток будет составлять время от одного деления до второго, т.е. совпадает с митотическим циклом. Несмотря на неограниченные спо-собности к делению и дифференпировке, стволовые клетки делятся очень редко и после завершения митоза пребывают в продленном С,-нериодс (иногда его называют Go-периодом). После деления стволовые клетки превращаются в полустволовые клетки, которые, наоборот, интенсивно делятся, восполняя клеточные потери. 2. Дифференцированные клетки. а) Необратимые постмитотические клетки. Такие клетки делятся митозом только в эмбриональном периоде, а затем после достижения популяцией необходимого объема полностью теряют способность к делению. Примером таких клеток являются нейроны, сердечные мышечные клетки. Жизненный цикл этих клеток состоит из следующих периодов: митотичес-кий цикл + детерминация (или определение пути дифференцировки) + диф-ференцировка (появление специфических черт строения клетки для выполнения специфической функции) + специализация ("обучение" функции, заключительные этапы дифференцировки) + период активного функционирования + старение + смерть клетки. б) Обратимые постмитотические клетки. Эти клетки (например, клетки печени) характеризуются тем, что могут выходить из митотического цикла и переходить в состояние G0, или покоя. При этом они имеют возможность для двух путей своего развития: или возвратиться в митотический цикл и делиться, или необратимо дифференцироваться и приступить к функции. Такие клетки являются резервом ткани. ТИПЫ КЛЕТОЧНЫХ ПОПУЛЯЦИЙ (СООБЩЕСТВ). МЕХАНИЗМЫ РЕГУЛЯЦИИ ГОМЕОСТАЗА В РАЗЛИЧНЫХ ТИПАХ КЛЕТОЧНЫХ ПОПУЛЯЦИЙ В многоклеточном организме численность любой клеточной популяции жестко регулируется. Механизмы тканевого гомеостаза сложны и многогранны. На их полюсах находятся митоз и аноптоз (запрограммированная клеточная гибель, см. ниже). Как отмечалось, существует три вида соматических клеток в зависимости от их жизненного цикла: стволовые; постмитотически обратимые; постмитотически необратимые. Из сочетания этих клеточных типов формируются клеточные популяции (например, ткани). По соотношению в клеточных популяциях клеток с различными клеточными циклами французский ученый К. Лсблоп разделил все клеточные системы (ткани и органы) па три большие группы. 1. Статические (стационарные) клеточные популяции. К этой группе он отнес такие клеточные популяции, в которых в зрелом состоянии имеются только дифференцированные клетки и полностью отсутствуют стволовые клетки. К таким клеточным популяциям относятся нервная и сердечная мышечная ткани. В них клетки в процессе дифференцировки необратимо к'ряют способность к делению, и общее число клеток не может увеличиваться. Напротив, с течением времени определенная часть клеток погибает путем амонтоза, а при старении организма этот процесс усиливается. 2. Растущие (увеличивающиеся в размерах) клеточные популяции. К :>той группе относятся клеточные популяции с очень низким в норме темпом пролиферативных процессов. Одновременно и потеря клеток очень низка. Такие популяции содержат: 1) очень незначительное количество стволовых клеток (по мнению некоторых авторов, они вообще отсутствуют); 2) дифференцированные клетки; 3) покоящиеся клетки. Примером такого типа популяций может служить паренхима печени, почек, щитовидной железы. В эмбриогенезе популяция гепатоцитов размножается с большой скоростью, в начале постнаталыгого развития число активно размножающихся гепатоцитов снижается, а продолжительность митотического цикла возрастает. У взрослых животных количество их падает до очень незначительных величин. Вместе с тем, при определенных ситуациях (удаление части органа) покоящиеся клетки быстро возвращаются в митотический цикл и, размножаясь, восстанавливают численность клеточной популяции. 3. Обновляющиеся популяции клеток. В этих популяциях достаточно интенсивное воспроизводство клеток уравновешивается такой же интенсивной их потерей за счет апоптоза. Такие популяции состоят из: 1) относительно небольшой фракции стволовых клеток, которые, делясь и дифференцируясь в зрелые клетки, поддерживают численность клеточной популяции. Эти клетки большую часть своего жизненного цикла проводят в состоянии продленного С,-периода. 2) постмитотически необратимых клеток, выполняющих основные функции популяции. В зависимости от типа клеточной популяции различны и механизмы поддержания ее гомеостаза. 1. В статических клеточных популяциях регуляторные механизмы направлены на регуляцию апоптоза. 2. В растущих клеточных популяциях регуляторные механизмы могут быть направлены на: 1) изменение процесса входа или выхода клеток в(из) состояние(я) покоя; 3) изменение длительности митотического цикла клеток; 2) изменение скорости дифференцировки клеток; 4) изменение интенсивности апоптоза клеток. 3. В обновляющихся клеточных популяциях регуляция может быть приложена к: 1) выходу или входу стволовой клетки из(в) продленного(ый) Go-период; 2) изменению длительности митотического цикла стволовых клеток; 3) изменению интенсивности апоптотической гибели клеток, которая в нормальных условиях достаточно высока. В последнем случае численность клеточной популяции будет зависеть от соотношения митотической активности и апоптотической гибели клеток: а) при уравновешивании их популяция находится в стационарном состоянии; б) при преобладании пролиферативных процессов отмечается прирост клеток, гипертрофия ткани, адаптация к вредным факторам; в) при преобладании апоптотической гибели наблюдается убыль клеточной популяции, что может иметь место при возвращении ткани после гипертрофии к исходному состоянию, атрофии ткани (малокровие, язвы и т.д.). ОБЩИЕ ЧЕРТЫ ПОКОЯЩИХСЯ КЛЕТОК (G0-W1ETOK) В покоящихся клетках существует ряд механизмов, поддерживающих макромолекулы в состоянии равновесия, необходимого для жизнедеятельности, и предотвращающих их гибель в неблагоприятных условиях. Благодаря этому покоящиеся клетки могут пребывать в состоянии покоя неограниченно долго, при этом в определенной мере выполняя некоторые специфические функции (в первую очередь это относится к клеткам, вышедшим в состояние покоя после С2-периода). В состоянии покоя может происходить репарация поврежденной ДНК. Для покоящихся клеток характерны следующие признаки: 1. По размерам они меньше пролиферирующих клеток. 2. В них более, чем в делящихся клетках, конденсирован хроматин. 3. В этих клетках низкий уровень синтеза ДНК. 4. Снижено содержание РНК при одновременном повышении процессов ее синтеза и распада. 5. Снижена проницаемость цитолеммы. 6. Снижена интенсивность метаболизма и дыхательная активность. ВНУТРИКЛЕТОЧНЫЕ, МЕЖКЛЕТОЧНЫЕ И ОРГАНИЗМЕННЫЕ МЕХАНИЗМЫ РЕГУЛЯЦИИ ДЕЛЕНИЯ КЛЕТОК. ВЛИЯНИЕ РАДИАЦИИ НА ДЕЛЕНИЕ КЛЕТОК Регуляция деления клеток осуществляется на разных уровнях. 1. ВНУТРИКЛЕТОЧНЫЙ УРОВЕНЬ. а) ГЕНОМНО-ЯДЕРНЫЙ УРОВЕНЬ. Осуществляется путем действия генов, ответственных за митоз. В клеточном геноме имеются "ранние" и "поздние" гены пролиферативного ответа. "Ранними" генами являются гены fos и туе, "поздними" — ras, myb-гены. Продуцируемые этими генами белки стимулируют вступление клетки в митоз и увеличивают его скорость. б) Цитоплазма может влиять на ядро с помощью различных растворимых факторов, так называемых триггерных белков, которые подавляют или активируют митоз. в) Компоненты цитолеммы (белки и гликопротеины) клетки могут оказывать регулирующее влияние на ее митотическую активность. Очевидно, они участвуют в таком феномене, как контактное торможение размножения, но могут выступать в роли самостоятельного регуляторного механизма. 2. МЕЖКЛЕТОЧНЫЙ УРОВЕНЬ регуляции митоза: а) Эффект контактного торможения размножения. Заключается в том, что при тесном контакте мембран двух клеток их деление подавляется. Это влияние опосредуется через плазмолемму, цитоскелет и передается на ядро. Эффект контактного торможения размножения объясняет, почему в культуре ткани клетки растут только в виде монослоя. Раковые же клетки не обладают этим эффектом, они растут беспорядочно, нарушая монослой. Аналогичным образом они ведут себя in vivo. б) Кейлонная регуляция. Кейлоны — вещества белковой природы. Они вырабатываются дифференцированными клетками, но способны действовать на стволовые клетки, подавляя их митоз. Кроме кейлонов, клетки выделяют еще специфические факторы роста, индукторы, которые стимулируют митоз. За счет кейлонов и индукторов регулируется постоянство клеточного состава тканей. 3. На ОРГАНИЗМЕННОМ УРОВНЕ регуляция деления идет за счет нервной, эндокринной, иммунной систем. Нервная система может как подавлять, так и стимулировать деление клеток в тканях-мишенях. Клетками эндокринной и иммунной системы вырабатывается большое количество ростовых факторов, которые стимулируют клеточное деление: эпидер-мальный, тромбоцитарный факторы роста, фактор роста нервов, фибробластов, инсулиноподобные факторы и др. Некоторые гормоны стимулируют, другие — подавляют размножение клеток. К гормонам-стимуляторам митотической активности клеток относят гормон роста, инсулин, тиреоидные гормоны и др. Подавляют размножение клеток глюкокор-тикоиды (гормоны коры надпочечников). Один и тот же гормон может оказывать на клетки разных органов различное влияние. Например, половые гормоны стимулируют деление клеток в половых органах, но подавляют его в зонах роста костей. Глюкокортикоиды подавляют деление клеток в большинстве органов, но могут стимулировать его в печени. Митоз является достаточно чувствительным к действию вредных внешних факторов процессом. Он может подавляться действием различных химических веществ, в том числе и лекарственных препаратов. Вещества, деполимеризующие микротрубочки веретена деления (например, колхицин), останавливают митоз в метафазе. Это используется для получения препаратов так называемых метафазных хромосом, которые наиболее удобны для изучения их строения. Эти же вещества используются для лечения опухолей. Радиация оказывает повреждающее влияние на митоз. Мито-тические хромосомы изменяют форму, возникают их разрывы, иногда с последующим неправильным соединением фрагментов. Порой отдельные хромосомы исчезают вообще. Наблюдаются аномалии веретена деления. Оно может иметь не два, а три полюса. В некоторых случаях хромосомы делятся, а деления ядра не происходит. В результате возникают большие ядра с полиплоидным набором хромосом. Если поражение радиацией сильное, то клетка теряет способность к делению. МЕХАНИЗМЫ КЛЕТОЧНОЙ РЕГЕНЕРАЦИИ Способность клетки или ткани восстанавливать утраченные части называется регенерацией. В зависимости от уровня ее реализации регенерация подразделяется на внутриклеточную регенерацию и регенерацию на клеточном уровне. Внутриклеточная регенерация — восстановление старых, разрушившихся органелл клетки, например, митохондрий, а также поврежденных частей клетки. В зависимости от назначения регенераторного процесса регенерация подразделяется на физиологическую и репаративную (посттравматическую) регенерацию. Физиологическая регенерация — это восстановление старых, подлежащих замене компонентов клетки или целых клеток. Репаративная регенерация — восстановление клеток после повреждения. После повреждения наряду с регенераторным процессом, как правило, происходят и компенсаторные, адаптивные изменения в клетке, направленные на уменьшение последствий возможного повторного повреждения. Если в результате количество органелл увеличивается, то это явление называется гиперплазией органелл. Если количество органелл остается тем же, но увеличиваются их размеры, то это явление называется гипертрофией органелл. Может наблюдаться сочетание гипертрофии и гиперплазии органелл. В результате указанных изменений клетка увеличивается в размерах (гипертрофия клетки) и становится менее чувствительной к действию вредных факторов. Клеточная регенерация — это регенерация ткани за счет увеличения числа клеток путем митоза. Подробнее о регенерации — в общей гистологии. РЕАКТИВНЫЕ ИЗМЕНЕНИЯ КЛЕТОК. ПОНЯТИЕ О ГИПЕРПЛАЗИИ И ГИПЕРТРОФИИ КЛЕТОК. СМЕРТЬ КЛЕТОК. НЕКРОЗ Реактивные изменения клеток — изменения структуры и функции клеток под воздействием внешних факторов. Если внешний фактор не вызы-иает гибели клеток, то в клетках происходят компенсаторные изменения, направленные на уменьшение последствий вредного воздействия внешнего фактора. Эти изменения могут быть следующими: 1. Внешний фактор активирует деление клеток. При этом клеток становится больше, и в результате распределения фактора на большее количество клеток они легче переносят его воздействие. 2. Внешний фактор воздействует на клетку, которая не может делиться. При этом в клетке активируется синтез белка, органелл, происходит их гиперплазия и гипертрофия. В результате клетка увеличивается в размерах и становится менее чувствительной к действию фактора. В первом случае мы имеем дело с гиперплазией клеток, во втором — с их гипертрофией. 3. Внешний фактор может привести к образованию полиплоидных и двуядерных клеток. Такие клетки больше по размерам, функционально более активны и менее чувствительны к повреждающему фактору. 4. Внешний фактор может вызвать усиление метаболизма и функциональной активности клеток. Это один из вариантов реакции клеток на внешний раздражитель. Может происходить увеличение клеточной поверхности и усложнение ее формы, что в определенной степени ведет к снижению интенсивности внешнего воздействия на единицу площади клетки. Как проявление реакции на внешние стимулы может активироваться фагоцитоз клеток, особенно тех, у которых он является основной функцией. Может возрастать двигательная активность клеток. Мышечные клетки на стимуляцию ответят сокращением, нервные — нервным импульсом, секреторные — усилением выработки и выделения секрета и т.д. 5. Любые достаточно сильные внешние воздействия вызывают в клетках стрессорные реакции, протекающие стереотипно. При этом происходит активация определенных генов, обеспечивающих синтез специальных защитных белков при одновременной блокаде других синтетических процессов. Эти защитные белки названы белками теплового шока (БТШ), поскольку первоначально были обнаружены в клетках при воздействии на них высокой температуры. БТШ носят универсальный характер, обладают собственной повышенной устойчивостью и одновременно предотвращают повреждение (агрегацию, коагуляцию) других клеточных белков, способствуют расщеплению возникших патологических белковых конгломератов. 6. При воздействии на клетку запредельных факторов она подвергается разрушению — некрозу. Обычно некроз захватывает целые группы клеток. Морфологические изменения при этом касаются и ядра, и цитоплазмы. В ядре могут происходить такие изменения. Под действием активированного лизосомалыгаго фермента ДНКазы ядерная ДНК расщепляется на фрагменты различной длины, что ведет к изменению расположения хроматина: он скапливается в виде крупных глыбок под кариолеммой. В дальнейшем с ядром могут происходить следующие изменения: — КАРИОПИКНОЗ — сморщивание ядра, уменьшение его в размерах до полного исчезновения; — КАРИОЛИЗИС — растворение ядра с постепенным исчезновением в нем всех структур. Ядро приобретает вид бесструктурного пузырька; — КАРИОРЕКСИС — разрыв ядра на отдельные фрагменты, которые затем разрушаются. Лишенная в результате одного из этих трех процессов ядра клетка становится нежизнеспособной и постепенно гибнет. В цитоплазме происходит дегенерация органелл. Цистерны ЭПС расширяются, гранулярная ЭПС полностью лишается рибосом. Матрикс митохондрий просветляется, расширяется межмембранное пространство, в дальнейшем разрушаются кристы; в конечном итоге мембраны митохондрий разрываются и митохондрии разрушаются. Повреждаются мембраны лизосом, ферменты которых выходят в цитозоль и участвуют в разрушении структур .цитоплазмы. Повреждение клеточных мембран связано с накоплением в гиалоплазме клетки кальция, который активирует связанные с мембранами ферменты фосфолипазы. В цитоплазме идет образование вакуолей — вакуольная дистрофия, накопление нетипичных белковых или жировых включений — белковая, жировая дистрофия. Активация лизосом приводит к аутолизу клетки и фагоцитозу ее макрофагами. ГЕНЕТИЧЕСКИ ЗАПРОГРАММИРОВАННАЯ КЛЕТОЧНАЯ ГИБЕЛЬ (АПОПТОЗ) Апоптоз часто называют физиологической, альтруистической гибелью клетки (в отличие от некроза, представляющего собой патологическую гибель клетки, смерть клетки от "несчастного случая"). Термин "апоптоз" (от греч. apoptosis — листопад) предложил в 1971 году Г. Керр, основываясь на внешнем сходстве апоптозных клеток с опадающими листьями: апоптозная клетка сморщивается и как бы выпадает из общего контекста ткани. Апоптоз является противоположностью митоза и генетически опосредован. В геноме каждой клетки наряду с генами пролиферации имеются гены апоптоза. Один из наиболее изученных механизмов индукции апоп-тоза — экспрессия гена Fas/Apo-1 (CD95). Находясь на противоположиых полюсах жизненного цикла клеток, митоз и апоптоз осуществляют регуляцию тканевого гомеостаза. Интересно, что одни и те же факторы в зависимости от конкретной ситуации могут выступать индукторами (ингибиторами) как митоза, так и апоптоза. МЕХАНИЗМ АПОПТОЗА Апоптоз индуцируется через рецепторы клеточной оболочки. Общую схему опосредования апоптоза можно представить следующим образом: 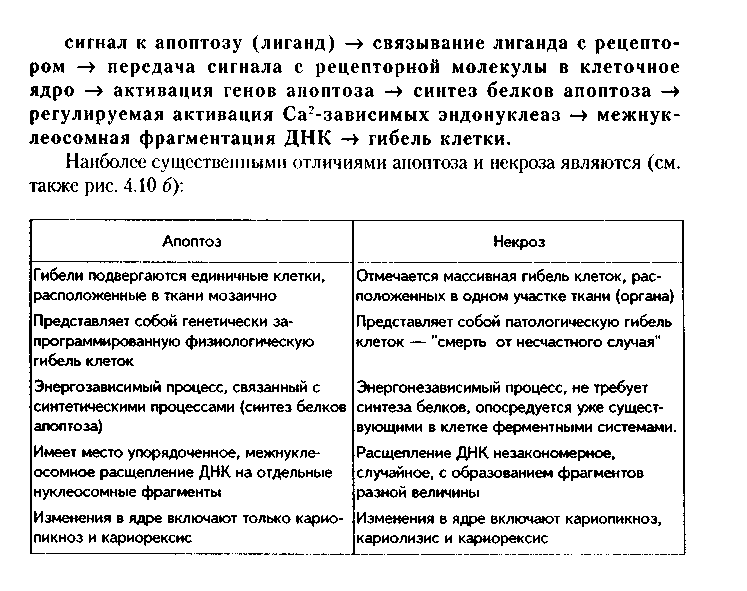 МОРФОЛОГИЯ АПОПТОЗА Изменения ядра. В результате правильной межнуклеосомной фрагментации ДНК происходит закономерная "упаковка" хроматина в ядре в виде своеобразных полулуний под кариолеммой. Ядро сильно уплотняется, иногда с зазубринами (рис. 4.10 а). В дальнейшем ядро распадается на несколько частей, окруженных мембраной (проявления кариопикноза и ка-риорексиса, но не кариолизиса). Изменения в цитоплазме. В результате прогрессивного активного распада клеточных органелл цитоплазма сжимается и уплотняется. В ней появляются оксифильные включения. Из-за сжатия цитоплазмы клетка оказывается окруженной своеобразным светлым бесструктурным ободком. Изменения клеточной поверхности. На поверхности клетки появляются многочисленные выпячивания и углубления (блеббинг). Выпячивания могут содержать неразрушенные органеллы и остатки ядра. В последующем эти выпячивания отшнуровываются, и клетка распадается на окруженные мембраной фрагменты — апоптозные тела, которые фагоцитируются местными макрофагами. РЕГУЛЯЦИЯ АПОПТОЗА Апоптоз, являясь одним и.ч важнейших факторов регуляции тканевого гомсостаза, так же, как и митоз, подвергается жесткой регуляции на разных уровнях. ' 3 1 Геномно-ядерный уровень.Наиболее изученным механизмом индукции апоптоза являет-ся экспрессия гена Fas/APO-1. Этот ген продуцирует специальный рецептор на клеточной поверхности, АРО-1, возбуждение которого запускает программу клеточного суицида. Одним из специфически связывающихся с рецептором АРО-1 факторов, запускающим апоптоз, является фактор некроза опухолей, продуцируемый макрофагами. На молекулярном уровне задействованы различные цитокины, ростовые и апоптозиндуцирую-щие факторы. 2. На межклеточном и тканевом уровне регуляция апоптоза осуществляется различными популяциями клеток. Например, клетки Лангерганса эпидермиса могут запускать апоптоз кератиноцитов. Подобная роль, во всяком случае, установлена для клеток Лагерганса многослойного эпителия шейки матки. Индуцировать апоптоз могут лимфоциты и, возможно, другие клетки. 3. Организменный уровень, а) Иммунная регуляция. Различные клетки иммунной системы и иммунные медиато-ры способны индуцировать аггаптоз. Доказан антителоопосредованный апоптоз. Он воз}1икает при действии антител на клеточные рецепторы. б) Гормональная регуляция. Одним из индукторов апоптоза являются стероидные гормоны. Клетки, лишенные рецепторов к стероидам, не подвергаются активной гибели. Это отмечается, например, в лейкозных клетках. Механизм действия глюкокортикоидов при апоптозе заключается в стимуляции межнуклеосомной фрагментации ДНК при помощи нуклеаз. Из-за своего апоптозиндушгрующего действия глюкокортикоиды используются для лечения злокачественных опухолей. В других случаях аполтоз индуцируется не избытком, а недостатком гормона. Так, при снижении в крови уровня мужского полового гормона тестостерона и адренокортикотропного гормона происходит усиление апоптотической гибели клеток соответственно в предстательной железе и в коре надпочечников. Эффект кастрации при раке предстательной железы, очевидно, основан на активации апоптоза раковых клеток, лишенных стимуляторов в виде андрогенов. С другой стороны, женские половые гормоны и их синтетические аналоги также усиливают апоптоз раковых клеток в предстательной железе. Нервная регуляция. Нервная система также участвует в регуляции апоптоза. Установлено, что апоптотическую гибель клеток можно индуцировать путем денервации органа. 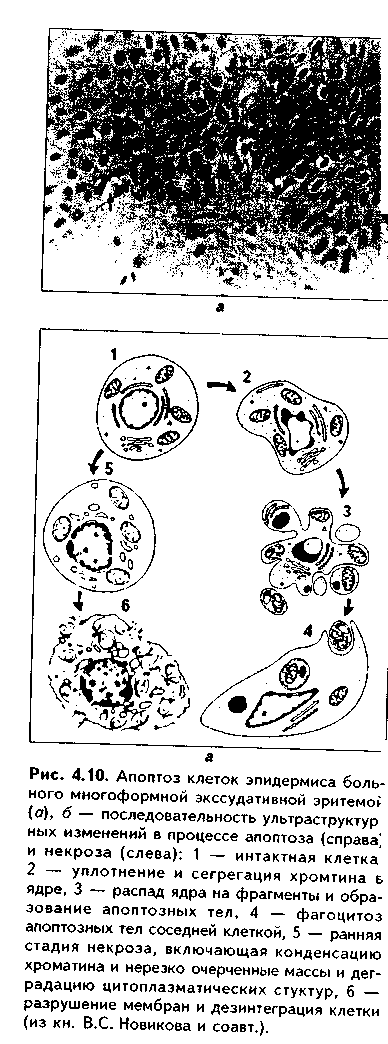 Ниже приведены вещества и факторы, ОБЩЕБИОЛОГИЧЕСКОЕ И МЕДИЦИНСКОЕ ЗНАЧЕНИЕ АПОПТОЗА 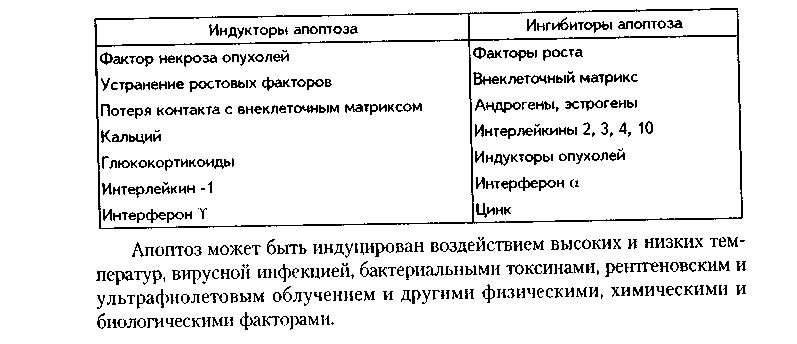 1. Апоптоз в эмбриогенезе. В ходе эмбриогенеза происходит не только рост, но и регрессия частей эмбриональных зачатков, тканей и органов: ин-волюция провизорных (временных) органов, появление просвета в полых органах. В целом эмбриональное развитие происходит с образованием большого избытка клеток, которые своевременно подвергаются апоптозу. Это создает запас клеточного материала и возможность последовательного развития. 2. Апоптоз стареющих клеток в тканях взрослого организма. При помощи апоптоза в организме ликвидируются старые клетки, которые замещаются новыми клетками, возникающими в результате деления стволовых клеток (обновляющиеся и растущие ткани). В статических тканях в результате постоянного апоптоза определенной части клеток количество клеточной популяции с возрастом снижается. 3. Апоптоз при инволюции зрелых тканей имеет место, например, в половых органах при старении, в матке и молочной железе при возвращении их размеров к норме после родов и грудного вскармливания и т.д. 4. Апоптоз в клетках иммунной системы имеет место при инволюции тимуса, в периферических органах иммунитета после иммунных реакций и т.д. 5. Апоптоз как реакция на слабые повреждения, слабые экстремальные внешние факторы (см. выше). 6. Апоптоз при атрофических, дегенеративных, инфекционных и онкологических заболеваниях. Существуют заболевания, связанные как с инги-бированием, так и с индукцией апоптоза. Ниже перечислены некоторые из этих заболеваний. 7. Апоптоз клеток в обновляющихся клеточных популяциях. В норме в таких клеточных популяциях в процессе деления стволовых клеток образуется большее, чем необходимо для поддержания гомеостаза, количество клеток. При этом излишние новообразованные клетки сразу же подвергаются апоптозу, тогда как другие вступают на путь дифференцировки. Такая кажущаяся расточительность является оправданной, т.к. создается материальная основа для адаптации ткани к внезапным функциональным нагрузкам на ткань. В этом случае в "лишних" клетках происходит замена программы апоптоза на программу дальнейшей дифференцировки, и количество функционирующих клеток быстро возрастает. 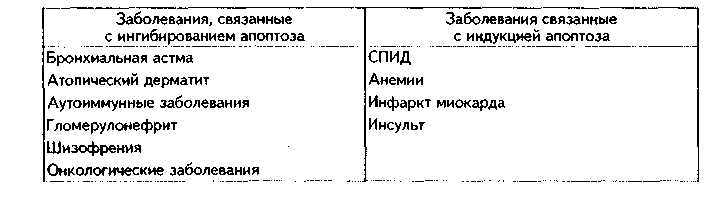 8. Аноптотичсская смерть мутировавших клеток, а также клеток, пораженных вирусом, способствует санации организма, освобождает его от таких клеток, в том числе и опухолевых. При онкологических заболеваниях подавляются факторы индукции апоптоза клеток. Такие клетки не могут погибнуть и вынуждены делиться приводя к развитию опухолей. Индукция апоптоза в этом случае лежит в основе лечебных мероприятий. С другой стороны, клетки иммунной системы осуществляют защиту организма от возникновения злокачественных новообразований путем индукции в опухолевых клетках апоптоза. 7 Апоптоз и проблема бессмертия. Мутации в генах апоптоза могут делать клетки бессмертными. Целенаправленные изменения этих генов возможное решение проблемы долголетия и бессмертия. |