зайко. Н. Н. Зайко Патологическая физиология Введение Предмет и задачи патологической физиологии Патологическая физиология есть наука, изучающая жизнедеятельность больного организма. Программа
 Скачать 7.32 Mb. Скачать 7.32 Mb.
|

|
Глава 7. Аллергия Аллергия (от греч, alios – иной, ergon – действую) – качественно измененная реакция организма на действие веществ антигенной природы, которая приводит к разнообразным нарушениям в организме – воспалению, спазму бронхиальной мышцы, некрозу, шоку и другим изменениям. Следовательно, аллергия – это комплекс нарушений, возникающих в организме при гуморальных и клеточных иммунных реакциях. Этиология. Причиной аллергии могут быть самые различные вещества с антигенными свойствами (аллергены), которые вызывают в организме иммунный ответ гуморального или клеточного типа. Аллергены разделяют на экзо- и эндогенные (рис. 7.1). Аллергенами могут быть полные антигены и неполные -гаптены. Неполные антигены вызывают аллергию несколькими путями: 1. соединяясь с макромолекулами организма, индуцируют выработку антител, специфичность которых направлена против гаптена, а не претив его носителя; 2. формируя антигенные комплексы с молекулами организма. При этом образовавшиеся антитела реагируют только с комплексом, а не с его компонентами. Аллергия может развиваться при воздействии на организм физических факторов и веществ, которые не являются антигенами, а только факторами, вызывающими появление антигенов. В данном случае физические факторы (тепло, холод, радиация) и химические вещества индуцируют в организме образование аллергенов из молекул организма путем демаскирования скрытых антигенных детерминант или образования новых антигенных детерминант в результате денатурации молекул. С выработанными антителами демаскирующий или денатурирующий агент не реагирует. Патогенез. Разнообразные по клиническим проявлениям аллергические реакции имеют общие патогенетические механизмы. Различают три стадии аллергических реакций: иммунную, биохимическую (патохимическую) и патофизиологическую, или стадию функциональных и структурных нарушений. Иммунная стадия аллергических реакций. Иммунная стадия начинается при первой встрече организма с аллергеном и заканчивается взаимодействием антитела с антигеном. В этот период происходит сенсибилизация организма, т. е. повышение чувствительности и приобретение способности реагировать на повторное введение антигена аллергической реакцией. Первое введение аллергена называется сенсибилизирующим, повторное же, которое непосредственно вызывает проявление аллергии, разрешающим. Сенсибилизация бывает активной и пассивной. Активная сенсибилизация развивается при иммунизации антигеном, когда в ответ включается собственная иммунная система. Механизмы активной сенсибилизации следующие: 1. Распознавание антигена, кооперация макрофагов с Т- и В-лимфоцитами, выработка плазматическими клетками гуморальных антител (иммуноглобулинов) или образование сенсибилизированных лимфоцитов (Т-эффекторов) и размножение лимфоцитов всех популяций. 2. Распределение антител (IgE, IgG) в организме и фиксация их на клетках-мишенях, которые сами антител не вырабатывают, в частности, на тканевых базофилах (тучных клетках), базофильных гранулоцитах, моноцитах, эозинофилах, а также на тромбоцитах, или взаимодействие иммуноглобулинов (IgG, IgM, IgA) либо Т-эффекторов с антигенами, если к моменту развития сенсибилизации они еще присутствуют в организме. На 7 – 14-й день после введения аллергена в сенсибилизирующей дозе организм приобретает к нему повышенную чувствительность. Пассивная сенсибилизация осуществляется в неиммунизированном организме при введении ему сыворотки крови, содержащей антитела, или клеточной взвеси с сенсибилизированными лимфоцитами, полученными от активно сенсибилизированного данным антигеном донора. При этом состояние повышенной чувствительности развивается через 18 – 24 ч. Это время необходимо для распределения антител в организме и фиксации их на клетках. На характер аллергической реакции влияют следующие особенности иммуноглобулинов (рис. 7.2). 1. Способность связывать комплемент, которая максимально выражена у IgM и умеренно у IgG. 2. Способность проникать в ткани, ограниченная у крупномолекулярных IgM, умеренно выраженная у IgG и сильно выраженная у IgE и IgD. 3. Способность сорбироваться на клетках тканей, сильно выраженная у IgE и свойственная некоторым субклассам IgG. 4. Способность преципитировать – наиболее выраженная у IgM и IgG. IgE обычно представляют собою непреципитирующие антитела. Однако реакция преципитации, агглютинации и флоккуляции зависят не только от класса иммуноглобулинов, но и от свойств антигена. Поэтому с некоторыми антигенами даже IgA могут давать реакцию преципитации. 5. Проникновение в секреты и слизь. Основным секреторным типом антител являются IgA. Однако в секреты и слизь могут транспортироваться также и IgG и даже IgM. 6. Способность проникать через плаценту, которая играет важную роль, с одной стороны, в индукции иммунитета у плода, а с другой – в возникновении иммунного конфликта между матерью и плодом и развитии аллергии у плода и новорожденного. Такой способностью у человека обладают преимущественно IgG. Кумбс и Джелл (1968) выделили следующие типы аллергических реакций: 1. Тип I – реагиновый (анафилактический). Антитела сорбированы на клетке, а антигены поступают извне. Комплексы антиген-антитело образуются на клетках, несущих антитела. В патогенезе реакций существенным является взаимодействие антигена с IgE и IgG, (реагинами), сорбированными на тканевых базофилах, и последующая дегрануляция этих клеток (рис. 7.3). Система комплемента при этом не активируется. К этому типу реакций относят анафилаксию общую и местную. Общая анафилаксия бывает при анафилактическом шоке. Местная анафилаксия подразделяется на. анафилаксию в коже (крапивница, феномен Овери) и анафилаксию в других органах (бронхиальная астма, сенная лихорадка). 2. Тип II – реакции цитолиза, или цитотоксические реакции. Антиген является компонентом клетки или сорбирован на ней, а антитело поступает в ткани. Аллергическая реакция начинается в результате прямого повреждающего действия антител на клетки; активации комплемента; активации субпопуляции В-киллеров; активации фагоцитоза. Активирующим фактором является комплекс антиген-антитело. К цитотоксическим аллергическим реакциям относится действие больших доз антиретикулярной цитотоксической сыворотки Богомольца (АЦС). 3. Тип III – реакции типа феномена Артюса или иммунных комплексов. Ни антиген, ни антитело при этом не являются компонентами клеток, и образование комплекса антиген-антитело происходит в крови и межклеточной жидкости. Роль преципитирующих антител выполняют IgM и IgG. Микропреципитаты сосредоточиваются вокруг сосудов и в сосудистой стенке. Это приводит к нарушению микроциркуляции и вторичному поражению ткани, вплоть до некроза. IgM, IgG – IgG, активируют комплемент, а посредством него – выработку других активных веществ, хемотаксис и фагоцитоз. Образуется лейкоцитарный инфильтрат – замедленный компонент феномена Артюса. 4. Тип IV – реакции замедленной гиперчувствительности (ГЗТ). Главная особенность реакций замедленного типа состоит в том, что с антигеном взаимодействуют Т-лимфоциты. Реакция замедленной гиперчувствительности не менее специфична по отношению к антигену, чем реакция с иммуноглобулинами, благодаря наличию у Т-лимфоцитов рецепторов, способных специфически взаимодействовать с антигеном. Этими рецепторами являются, вероятно, IgM, укороченные и встроенные в мембрану Т-лимфоцита, и антигены гистосовместимости (см. ниже). Однако в ткани, где происходит эта реакция, среди множества клеток, разрушающих антиген и ткань, обнаруживается только несколько процентов Т-лимфоцитов, способных специфически реагировать с антигеном. Данный факт стал понятен после открытия лимфокинов – особых веществ, выделяемых Т-лимфоцитами. Благодаря им иммунные Т-лимфоциты даже в небольшом количестве становятся организаторами разрушения антигена другими лейкоцитами крови (см. ниже). 5. Тип V – стимулирующие аллергические реакции. В результате действия антител на клетки, несущие антиген, происходит стимуляция функции этих клеток. Механизм стимуляции объясняется тем, что выработанные антитела могут специфически реагировать с рецепторами клетки, предназначенными для активирующих гормонов или медиаторов. К стимулирующему типу аллергических реакций относится аутоиммунный механизм базедовой болезни, приводящий к гиперфункции щитовидной железы. В зависимости от времени появления реакции после контакта с аллергеном различают также аллергические реакции немедленного типа (гиперчувствительность немедленного типа – ГНТ) иаллергические реакции замедленного типа (гиперчувствительность замедленного типа – ГЗТ) по классификации, предложенной R. A. Cooke (1930). В первом случае реакция развивается в течение 15 – 20 мин, во втором – через 1 – 2 сут. Эта классификация существует и в настоящее время, однако она не отображает всего многообразия проявлений аллергий в том числе патогенетических особенностей, лежащих в основе классификации Джелла и Кумбса. Особенности иммунной стадии реакций замедленного (клеточного) типа. Т-лимфоциты распознают антигенные детерминанты с высокой степенью специфичности с помощью рецепторов, в состав которых входит антиген главного комплекса гистосовместимости МНС (от англ. major Histocompatibility complex). Гены, кодирующие антигены МНС, располагаются у человека в 6-й хромосоме, имеется их 4 аллеля, каждый из генов встречается в генофонде во множестве (десятки) вариантов. Антигены МНС являются веществами, встроенными в мембраны клеток, в том числе, в мембраны лейкоцитов, поэтому их обозначают НLA-A, HLA-B, HLA-C, HLA-D (от англ. Human Leucocytes Antigen – антиген лейкоцитов человека). По участию в иммунных реакциях лимфоцитов вещества главного комплекса гистосовместимости МНС разделили на две группы: к группе HI относятся HLA-A, HLA-B, HLA-C, группа НII включает HLA-D. В состав рецепторов Т-киллеров входят вещества группы HI. В том же организме рецепторы Т-хелперов содержат вещества из группы НИ (аллель HLA-D). Установлено, что клетки организма встраивают чужеродные антигены в свою мембрану в вещество комплекса МНС, например, антигены вируса при заражении клетки. Т-лимфоцит может распознать чужеродный антиген, если это чужеродное вещество в клетке-носителе встроено в такой же антиген главного комплекса гистосовместимости, каким располагает сам Т-лимфоцит, т. е. происходит ассоциированное иммунное распознавание. Патохимическая стадия аллергических реакций I – III типа. Сущность биохимической стадии заключается в образовании или активации биологически активных веществ (БАВ), которое начинается уже с момента соединения антигена с антителом (рис. 7.4). При этом происходят следующие процессы. 1. Активация системы комплемента (кроме I типа). Активный комплемент обладает ферментной активностью, способностью разрушать мембраны микроорганизмов и тканевых клеток (см. выше), вызывая при этом освобождение новых БАВ, способностью активировать фагоцитоз, протеолитические ферменты крови, фактор Хагемана, дегрануляцию тканевых базофилов. Часть фракций комплемента СЗа и С5а не включаются в общий комплекс, а действуют как самостоятельные биологически активные вещества, которые вызывают: дегрануляцию базофилов крови и тканевых базофилов; у нейтрофилов – хемотаксис, адгезию к эндотелиоцитам сосудов, образование и освобождение лейкотриенов; у макрофагов – хемотаксис, секрецию гликолитических и протеолитических ферментов, продукцию интерлейкина 1; у лимфоцитов – различное селективное действие на разные субпопуляции лимфоцитов, модулируют рециркуляцию, пролиферацию и дифференцировку лимфоцитов, обработку лимфоцитами антигенов. Компоненты СЗа и С5а являются анафилотоксином, существование которого как медиатора анафилаксии предполагали ранее. СЗа и С5а вызывают увеличение проницаемости сосудов и сокращение гладкой мускулатуры. 2. Активация фактора Хагемана – XII фактора свертывания крови. Активированный фактор Хагемана в свою очередь активирует свертывающую систему крови, систему комплемента и протеолитические ферменты крови. 3. Активация протеолитических ферментов крови – трипсиногена, профибринолизина, калликреиногена. Биологическая активность этих ферментов проявляется в расщеплении белков на полипептиды, во взаимоактивации друг друга, в активации фактора Хагемана, системы комплемента, тканевых базофилов, в способности повреждать клетки тканей, разрушать с помощью фибринолизина фибрин. Калликреин, являясь ведущим звеном в кининовой системе крови, отщепляет от глобулина крови – кининогена – полипептиды, которые называются кининами. К ним относятся брадикинин – нонапептид (Н2 – Apr – Про – Про – Гли – Фе – Сер – Про – Фе – Apr – СООН) и каллидин – декапептид, который под действием трипсина превращается в брадикинин. Брадикинин вызывает повышение проницаемости сосудов, снижение тонуса и их расширение, спазм неисчерченной мышечной ткани некоторых органов, является медиатором боли. Каллидин менее активен, чем брадикинин. 4. Выделение в ткань из окончаний чувствительных нервов полипептида Р, являющегося одним из самых сильных медиаторов воспаления. Полипептид Р резко увеличивает проницаемость сосудов, активирует другие системы БАВ: вызывает дегрануляцию тканевых базофилов, активирует кининовую систему др. 5. Активация и освобождение протеолитических ферментов тканей – катепсинов и тканевой гиалуронидазы. 6. Дегрануляция тканевых базофилов, на которых сорбированы IgE, IgG, происходит при присоединении к иммуноглобулинам антигена. При этом выделяются две группы биологически активных веществ: 1. синтезируемые заранее (преформированные), связанные с гранулами – гистамин, гепарин, серотонин, фактор хемотаксиса эозинофилов, высокомолекулярный фактор хемотаксиса нейтрофилов, воспалительный фактор анафилаксии, различные ферменты (протеазы, кислые гидролазы и др.); 2. образуемые в процессе дегрануляции из компонентов мембраны – лейкотриены: С4 и D4 – медленно реагирующая субстанция МРС-А, лейкотриен В4, известный также как фактор хемотаксиса эозинофилов; простагландины, разнообразные по действию – активирующие и ингибирующие; фактор активации тромбоцитов (образующиеся при активации калликреин-кининовой системы – каллидин, брадикинин). Гистамин через рецепторы Н1 и серотонин, подобно брадикинину, повышают проницаемость сосудов, вызывают сокращение бронхиальной мышцы, кишок, матки; боль, зуд, жжение, шок и некроз, действуют на другие нервные рецепторы. В то же время действие гистамина на рецепторы типа Н2 вызывает противоположный эффект. Гепарин препятствует свертыванию крови, тормозит выработку антител, хемотаксис. Медленно реагирующая субстанция А вызывает постепенное, но длительное сокращение бронхиальной мышцы, что имеет важное значение в патогенезе бронхиальной астмы. Освобождение гистамина и серотонина происходит при распаде тромбоцитов и базофильных гранулоцитов. Липидные биологически активные вещества и перекисное окисление липидов. Известны 2 группы липидных БАВ, участвующих в аллергии: • производные фосфолипидов. К ним относятся производные 1-алкил-2-ацетилфосфатидилхолина, в частности фактор активации тромбоцитов; • продукты перекисного окисления производных арахидоновой кислоты – эйкозатетраеновой кислоты – эйкозаноиды. Перекисное окисление арахидоновой кислоты может идти двумя путями: липооксигеназным – под действием ферментов липооксигеназ образуются лейкотриены, сюда входят две группы: 1 – лейкотриены В; 2 – сульфолейкотриены, обозначаемые как лейкотриены С, D, Е. Сульфолейкотриены образуются при переносе цистеиновой сульфогруппы от глутатиона. Спазм бронхиол при бронхиальной астме вызывает медленно реагирующая субстанция А (МРС-А), которая является лейкотриеном D4. Второй путь окисления арахидоновой кислоты – циклооксигеназный – под действием ферментов циклооксигеназ образуются простагландины и тромбоксаны. Простагландины являются эндопероксидами, это ПГГ2, ПГН2, ПГФ2, ПГЕ2, ПГД2. Из тромбоксанов известен ТхА2. Эйкозаноиды обладают разнообразным биологическим действием: ПГФ2вызывает снижение тонуса сосудов, спазм неисчерченной мышечной ткани матки, бронхов, лизис тканевых базофилов и др. В то же время ПГ способствует накоплению цАМФ в клетках, тем самым вызывая расслабление бронхиальной мышцы снижение выделения из тканевых базофилов биологически активных веществ. Фактор активации тромбоцитов (ФАТ) вызывает бронхоспазм и в то же время расширяет сосуды с падением артериального давления, снижает коронарный кровоток. 7. Накопление продуктов разрушения клеток крови и тканей. 8. Распад лейкоцитов и освобождение лизосомальных факторов (см. раздел XII – "Воспаление"); изменение активности холинэстеразы и увеличение освобождения ацетилхолина; изменение содержания электролитов. Наблюдается повышение концентрации ионов калия и кальция, что приводит к изменению возбудимости тканей. 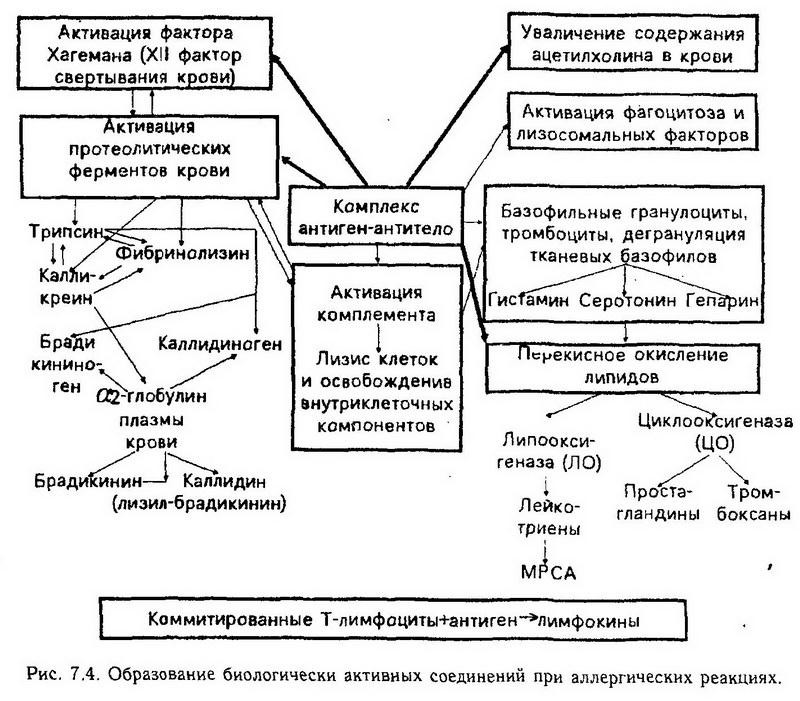 Все системы БАВ связаны между собою прежде всего механизмами взаимоактивации, но имеются и тормозные пути. При развитии аллергических реакций могут приобретать особое значение отдельные системы БАВ: 1. заболевания, зависимые от тканевых базофилов: анафилаксия, бронхиальная астма, крапивница; 2. зависимые от комплемента: васкулиты, пневмониты; 3. реакции цитотоксического типа, при которых особое значение имеет прямое повреждающее действие антител с активацией комплемента: васкулиты, пурпура (мелкие кровоизлияния), аутоиммунная гемолитическая анемия и др. Патохимическая стадия аллергических реакций замедленного типа. Реакция замедленной гиперчувствительности (IV тип) осуществляется при непосредственном контакте лимфоцита с антигеном. Если антиген является компонентом какой-нибудь клетки, то прикрепленные к этой клетке Т- или В-киллеры приводят к. гибели клетки-мишени. Киллер выделяет перфорин, который, подобно компонентам комплемента (см. рис. 6), встраивается в мембрану клетки или микроорганизма и полимеризуется, образуя в мембране сквозные каналы, что приводит к быстрой гибели клетки-мишени. При контакте с антигеном Т-лимфоциты вырабатывают лимфокины, которые представляют собой биологически активнее вещества. С помощью лимфокинов Т-лимфоциты управляют функцией других лейкоцитов. |