Ности. Связывание ангиотензина с рецепторами вызывает увеличение прогiииаемости
 Скачать 458.29 Kb. Скачать 458.29 Kb.
|

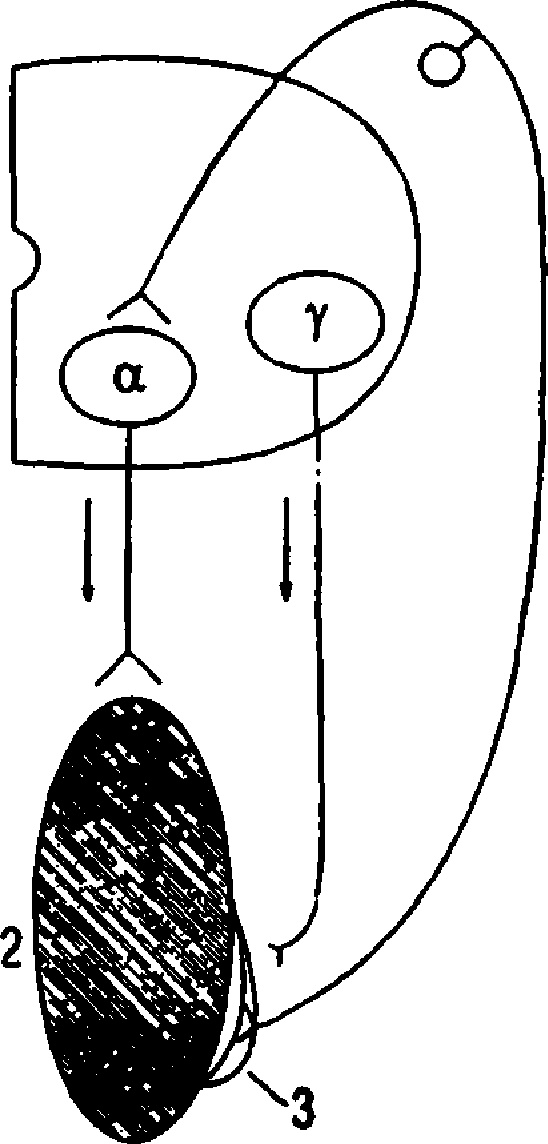 Рис. 8.4. Механизм регуляции тонуса мыши ниж их конечностей и развития гипертонуса в случае повреждения спинного мозга. 1 — полусегмент спинного мозга; 2 — скелетная мышка; 3 - мышечное веретено; стрелка вверх - афферентный путь; стрелка вниз - эфферентныс путы; а-мотонсйрон, иннервируюший скелстную мышку; у-мотонейрон, иннер-
нижних конечностей у человека в случае по-
Гипертонус имеет рефлекторную природу, он развивается вследствие афферентной им пульсаиии от мышечных рецепторов. Деаф ферентаиия, например, нижней конечности у лягушки ведет к исчезновению тонуса у нее (опыт бронжеста). 121 Имеется три фактора, обеспеч и ваюших афферентную импульсацию от мышечных ре цепторов по задним корешкам спинного мозга, которая ведет к возбуждению а-мото нейронов спинного мозга и сокращению ске летных мыши: 1) растяжение мышечных ре цепторов под влиянием сил гравитации ко нечности: 2) спонтанная активность мышеч ных рецепторов; 3) спонтанная активность у-мотонейронов, которая реализуется следу ющим образом. Возбуждение у-мотонейро нов вызывает возбуждение и сокращение ин нервируемых ими интрафузальных мышеч ных волокон, в результате чего увеличивается натяжение мышечного рецептора, поскольку концы его зафиксированы на скелетной мышце. Вследствие натяжения мышечного веретена раздражаются и возбуждаются аф ферентные окончания (проприореиепторы), импульсы от которых поступают к а-мото нейронам и возбуждают их. В свою очередь а-мотонейроны посылают импульсы к ске летной мышке и вызывают ее постоянное (тоническое) сокращение. Мы рассмотрели рефлекторную и провод никовую функции изолированного спинного мозга. В нормальных условиях деятельность спинного мозга контразируется вышележа шими отделами ЦНС посредством импульса иии ко всем его нервным элементам и полу чения обратной афферентаиии от всех орга нов и тканей. 8.2. СТВОЛ ГОЛОВНОГО МОЗГА Головной мозг состоит из конечного мозга (кора большого мозга, белое вещество, ба зальные ганглии), промежуточного, средне го, заднего (мост и мозжечок) и продолгова того мозга. Часть этих структур определяют понятиемствол мозга» (продолговатый мозг, мост и средний мозг), совместная дея тельность которых формирует основные стволовые функции, например, сложные цепные рефлексы, регуляцию мышечного то нуса и позы, восходящее влияние ретикуляр ной формации на конечный мозг. 8.2.1. ФУНКЦИИ СТВОЛА МОЗГА, РЕАЛИЗУЕМЫЕ ЯДРАМИ ЧЕРЕПНЫХ НЕРВОВ В стволе мозга находятся ядра 111—Х11 пар черепных нервов. Ядра глазодвигательного нерва (n. oculomotorius, 111) расположены в среднем мозге. Двигателыое ядро вызывает сокращение верхней, нижней, внутренней прямых, нижней косой мыши глаза и мышцы, поднимающей верхнее веко, участ вуя в глазодвигательных рефлексах. Добивоч иое (парасимпатическое) ядро, иннервируя сфинктер зрачка и ресничную мышцу, осу ществляет рефлексы сужения зрачка и акко модации глаза. Ядро блокового нерва (п. troehlearis, lV) на ходится в среднем мозге. Иннервируя верх нюю косую мышку, оно вызывает поворот глазного яблока вниз и кнаружи. Тройничный нерв (n. trigeminus, V) имеет двигательное и чувствительные ядра, лвигл тельное ядро расположено в мосту, иннерви рует жевательные мышцы и вызывает движе ния нижней челюсти (вверх, вниз, в стороны и вперед), а также напрягает мягкое небо и барабанную перепонку. Чувствительные ядра (среднемозговое, мостовое, спинальное) по лучают от кожи, слизистых оболочек, орга нов лица и головы тактильную, температур ную, висцеральную, проприоцептивную, бо левую импульсацию, входят в проводнико вый отдел соответствующих анализаторов и участвуют в различных рефлексах (например, жевательном, глотательном, чихательном). Ядро отводящего нерва (n. abducens, V1) расположено в мосту. Иннервирует наруж ную прямую мышку глаза, вызывает поворот его кнаружи. Ядра лицевого нерва (п. facialis, VII) нахо дятся в мосту. Двигательное ядро вызывает сокрашения мимической и вспомогательной жевательной мускулатуры, регулирует пере дачу звуковых колебаний в среднем ухе в ре зультате сокрашения стременной мышцы. Чувствительное ядро одиночного пути, и н -нервируя вкусовые луковицы передних 2/3 языка, анализирует вкусовую чувствитель ность, участвует в моторных и секреторных пишеварительных рефлексах. Верхнее слюно отделительное (парасимпатическое) ядро стимулирует выделение секретов подъязыч ной, подчелюстной слюнных и слезной желез. Ч увств ител ьн ы е ядра преддверно-улитко еого нерва (n. vestibuIocochIearis, V1 [ [) распо ложены в продолговатом мозге. Вестибуляр ные ядра, иннервируя рецепторы вестибуляр ного аппарата, участвуют в регуляции позы и равновесия тела (статические и статокинети ческие рефлексы), в вестибулоглазных и вес тибуловегетативных рефлексах, входят в про водниковый отдел вестибулярного анализа тора. Улитковые ядра, иннервируюшие слу ховые рецепторы, участвуют в слуховом ори ентировочном рефлексе, входят в проводни ковый отдел слухового анализатора. 122 Ядра языкоглоточного нерва (n. glosso pharyngeus, IX) расположены в продолгова том мозге. Двойное (двигательное) ядро вызы вает поднимание глотки и гортани, опуска ние мягкого неба и надгортанника при глота тельном рефлексе. Чувствительное ядро оди ночного пути получает вкусовую, тактильную, температурную, болевую и интероцептивную чувствительность от слизистой оболочки глотки, задней трети языка, барабанной по лости и каротидного тельца, входит в состав соответствующих анализаторов, участвует в рефлексах жевания, глотания, в секреторных и моторных пишеварительных рефлексах, в сосудистых и сердечных рефлексах (из каро тидного тельца). Нижнее слюноотделительное (парасимпатическое) ядро стимулирует секре цию околоушной слюнной железы. Ядра блуждающего нерва (n. vagus, Х) рас положены в продолговатом мозге. Двойное (двигательное) ядро, иннервируя мышцы неба, глотки, гортани, участвует в рефлексах глотания, рвоты, чиканья, кашля, в форми рова н и и голоса. Чувствительное ядро одиноч ного пути, иннервируя слизистую оболочку неба, корня языка, дыхательных путей, аор тальное тельце, органы шеи, грудной, брюш ной полостей, участвует в качестве аффе рентного звена в глотательном, жевательном, дыхательных, висиеральных рефлексах. Оно входит в проводниковый отдел интероцеп тивного, вкусового, тактильного, темпера турного и болевого анализаторов. Заднее (парасимпатическое) ядро, иннервируя серд це, гладкие мышцы и железы органов шеи, грудной и брюшной полостей, участвует в сердечных, легочных, бронхиальных, пише варительных рефлексах. Двигательное ядро добавочного нерва (n. accessorius, Х1) расположено в продолго ватом и спинном мозге. Иннервируя груди но-ключ ично-сосцевидную и трапециевид ную мышцы, оно вызывает наклон головы набок с поворотом лица в противоположную сторону, поднимание плечевого пояса вверх, приведение лопаток к позвоночнику. Двигательное ядро нодьязычного нерва (п . hypoglossus, XlI) расположено в продолгова том мозге; иннервируя мышцы языка, вызы вает его движение в рефлексах жевания, со сан ия, глотания, в осуществлении речи. Таким образом, с участием ядер черепных нервов реализуется сенсорная и рефлектор ная (соматическая и вегетативная) функции ствола мозга. Сенсорная функция связана с чувствитель ными ядрами черепных нервов, в которых импульсация с первых афферентных нейро нов переключается на вторые нейроны. В них осуществляется первичный анализ силы и качества (вида) раздражителей, обработанная информация передается в высшие афферент ные центры (например, таламус). Вкусовая чувствительность анализируется в ядрах ли цевого, языкоглоточного и блуждающего нервов. Чувствительность кожи и слизистых оболочек лица и головы (тактильная, темпе ратурная, болевая), а также мышечно-сустав- ная (проприоцептивная) чувствительность анализируется в ядрах тройничного нерва. И нтероiiептивная чувствительность органов головы, грудной и брюшной полостей оцени вается в ядрах блуждающего и языкоглоточ ного нервов. Слуховая и вестибулярная чув- ствительность анализируется в соответствую щих ядрах преддверно-улиткового нерва. С участием ствола мозга осушествляются сложные соматические рефлексы, в каждом из которых участвуют ядра несколь ких черепных нервов. Например, глазодвига тельные рефлексы имеют центры, функцио нально объединяюшие чувствительные ядра тройничного, преддверно-улиткового нервов, бугорки четверохолмия, двигательные ядра глазодвигательного, блокового и отводящего нервов, координация деятельности которых осуществляется ретикулярной формацией ствола мозга, а также мозжечком и корой большого мозга. В результате этих рефлексов осуществляются различные движения глаз. Рефлексы мимических мышц возникают с рецепторов тройничного нерва, а при генерализованных реакциях - с рецеп торов туловища, конечностей, внутренних органов. Афферентная импульсация из чув ствительных ядер V нерва переключается в двигательное ядро лицевого нерва, что вызы вает сокращение мимических мышц. К этим рефлексам относятся, например, сокращение круговой мышцы глаза (мигательный реф лекс) при ударе неврологическим молоточ ком по коже лица в области надбровной дуги или сокращение круговой мышцы рта при ударе по периоральной области. Рефлексы я з ы к а осушествляются со всех видов рецепторов языка. Афферент ная импульсация идет в чувствительные ядра тройничного, лицевого, языкоглоточного и блуждающего нервов и переключается в дви гательное ядро подъязычного нерва, имею шего группы нейронов, иннервируюшие раз личные мышцы языка. Сокращение этих мышц вызывает движение языка в различных направлениях. деятельность мотонейронов мышц языка координируется ретикулярной формацией ствола и корой большого мозга. Рефлексы языка входят в состав более слож ных (цепных) рефлексов - жевания, со сания, глотания, рвоты. Вегетативные рефлексы ствола головного мозга, осуществляемые через пара-симпатические ядра черепных нервов (III, VII, IX, Х), рассмотрены в соответствующих главах. 8.2.2. РЕТИКУЛЯРНАЯ ФОРМАЦИЯ СТВОЛА МОЗГА Ретикулярная формация (РФ) образована со вокупностью нейронов, расположенных в его центральных отделах как диффузно, так и в виде ядер. 123 Структурные особенности РФ. Нейроны РФ имеют длинные маловетвящиеся дендриты и хорошо ветвящиеся аксоны, кото рые часто образуют Т-образное ветвление: одна из ветвей аксона имеет нисходящее, а вторая - восходящее направления. Ветви нейронов под микроскопом образуют сеточку (ретикулум), с чем и связано название данной структуры мозга, предложенное О. дейтерсом (1865). Функциональные особенности ретикуляр ных нейронов.
Главные ядерные структуры и их функции. Латеральное ретикулярное ядро продолговато го мозга входит в центр кровообрашения, способствуя повышению АД и частоты сер дечных сокращений, оказывает активирую-шее влияние на кору, передает на мозжечок спинальные влияния, участвует в регуляции быстрых движений глаз, движений челюсти и языка в рефлексах сосания, жевания и глота ния. Ретикулярное гигантоклетоиное ядро продолговатого мозга повышает тонус мыши-сгибателей, входит в противоболевую систему мозга, в дыхательный центр и центр кровообрашения, оказывает активируюшее влияние на кору мозга. Ларамедианкое рети кулярное ядро продолговатого мозга входит в состав центра глотания, участвует в регуля ции содружественных движений глаз, в осу шествлении сердечно-сосудистых рефлексов, передает на мозжечок влияние коры мозга. Ретикулярное вентральное ядро продолговато го мозга входит в депрессорную зону центра кровообрашения, оказывает активируюшее влияние на кору мозга. Ретикулярное мелко-клеточное ядро продолговатого мозга входит в экспираторную область дыхательного центра. Каудальное ретикулярное ядро моста входит в депрессорную зону сосудодвигательного центра, повышает тонус мышц-разгибателей. Ретикулярное ядро покрышки моста (Бехтере- Повышение тонуса раз- гибателей и с н и ж е н и е тонуса сгмбателей ва) передает корковые и спинальные влияния на мозжечок. Оральное ретикулярное ядро моста входит в депрессорную зону центра кровообращения, повышает тонус мышц- разгибателей, участвует в регуляции частоты дыхания. Оральное интерстициальное ядро среднего мозга участвует в регуляции пово рота верхней части туловища и вращательных движений. Ретикулярная формация имеет связи с многочисленными структурами ЦНС. Аффе |