Ности. Связывание ангиотензина с рецепторами вызывает увеличение прогiииаемости
 Скачать 458.29 Kb. Скачать 458.29 Kb.
|

Тонические сгибательные (как и разгиба тельные) рефлексы конечностей возникают при длительном растяжении мыши и возбуж дении проприорецепторов, их главное назна чение — поддержание позы. Тоническое со кращение скелетных мышц является фоно вым для всех двигательных актов, осущест вляемых с помощью фазных сокращений мышц, обеспечивает сохрагiение позы. 2. Разгибательные рефлексы конечностей, как и сгибательные, бывают фазными и то ническими, возникают с проприорецепторов мышц-разгибателей, являются моносинапти ческими. Фазные рефлексы возникаютв ответ на однократное раздражение мышеч-
Тонические разгибательные р е ф л е к с ы представляют собой длитель ное сокращение мыши-разгибателей при длительном растяжении их сухожилий. Их роль — поддержание позы. В положении стоя тоническое сокращение мышц-разгибателей предотвращает сгибание нижних конечнос тей и обеспечивает сохранение вертикальной естественной позы. Тоническое сокращен не мышц спины удерживает туловище в верти кальном положении, обеспечивая осанку че ловека. Тонические рефлексы на растяжение мышц (сгибателей и разгибателей) называют таi9же миотатическими. /3. Позные рефлексы конечностей — пере распределение мышечного тонуса, возникаю щее при изменении положения тела или от дельных его пастей. Позные рефлексы осу ществляются с участием различных отделов цНС. На уровне спинного мозга замыкаются шейные позные рефлексы, наличие которых установил голландский физиолог Р.Магнус (1924) в специальных опытах на кошке. Име ется две разновидности этих рефлексов —возникающие при наклоне и при поворотах головы. При наклоне головы вниз (кпереди) уве личивается тонус мыши—сгибателей перед них конечностей и тонус мышц -- разгибате лей задних конечностей, в результате чего передние конечности сгибаются, а задние разгибаются. При наклоне головы вверх (кзади) возникают противоположные реак ции: передние конечности разгибаются вследствие увеличения тонуса их мышц-раз гибателей, а задние конечности сгибаются вследствие повышения тонуса их мыши-сги бателей. Эти рефлексы возникают с пропри ореиепторов мышц шеи и фасиий, покрыва ющих шейный отдел позвоночника. В усло виях естественного поведения животных они увеличивают шансы достать пищу, находя щуюся выше или ниже головы. Вторая группа шейных позных рефлексов возникает с тех же рецепторов, но только при поворотах или наклонах головы вправо или влево. При этом повышается тонус мышц разгибателей обеих конечностей на стороне, куда повернута (наклонена) голова, и повы шается тонус мышц-сгибателей на противо положной стороне. Рефлекс направлен на со хранение позы, которая может быть наруше на вследствие изменения положения центра тяжести после поворота (наклона) головы. центр тяжести смещается в сторону поворота головы — именно на этой стороне повышает ся тонус мышц--разгибателей обеих конеч ностей. 118 4. Ритмические рефлексы — многократное повторное сгибание и разгибание конечнос тей (рефлексы потирания, чесания и шага тельный рефлекс). Рефлекс потирания за ключается в том, что после смазывания рас твором кислоты кожи бедра спинальная ля гушка (лягушка, у которой удален головной мозг) многократно потирает этот участок - пытается освободиться от раздражителя. Сла бое раздражение кожи боковой поверхности тела у собаки вызывает почесывание этого участка задней конечностью. Чесательный рефлекс — аналог рефлекса потирания у ля гушки и шагательный рефлекс изучили у подвешенной с помощью лямок в станке спинальной собаки в стадии хирургического наркоза. Б. Брюшные рефлексы — верхний, средний и нижний. Все они вызываются штриховым раздражением кожи живота, выражаются в сокращении соответствующих участков мус кулатуры стенки живота; это защитные реф лексы. Для вызова верхнего брюшного реф лекса раздражение наносят параллельно нижним ребрам непосредственно под ними, дуга рефлекса замыкается на уровне грудных сегментов спинного мозга (Th111—Тнiх). Средний брюшной рефлекс вызывают раз дражением на уровне пупка (горизонтально), дуга рефлекса замыкается на уровне Тнiх Th, |
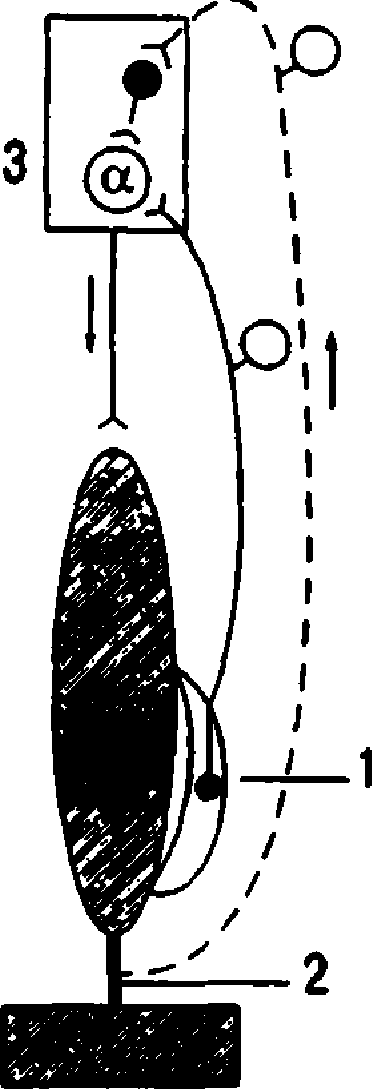
| мозят нейроны своего центра и возбуждают нейроны центра-антагониста (у мыши-сгиба телей это возбуждение выражено слабее). Элемент шагательного рефлекса - пооче редное сокращение и расслабление скелет ной мышцы под влиянием импульсов от про приореиепторов, поступающих в свой центр, осуществляется следующим образом. Когда мышка (сгибатель или разгибатель) расслаблена и удлинена, возбуждаются мы шечные веретена, импульсы от них поступа ют к своим а-мотонейронам спинного мозга и возбуждают их (см. рис. 8.2, А). Далее а мотонейроны. посылают импульсы к этой же скелетной мышце, что ведет к ее сокраще нию. Как только мышка сократилась (см. рис. 8.2, Б), возбуждение мышечных веретен прекращается или сильно ослабляется (они уже не растянуты), начинают возбуждаться сухожильные рецепторы. Импульсы от пос ледних поступают тоже в первую очередь в свой центр в спинном мозге, но к тормозным клеткам Реншоу. Возбуждение тормозных клеток вызывает торможение а-мотонейро нов этой же скелетной мышцы, вследствие чего она расслабляется. Однако ее расслабле ние (удлинение) ведет снова к возбуждению мышечных веретен и а-мотонейронов —мышца вновь сокращается. Вследствие ее со краинения возбуждаются сухожильные рецеп торы и тормозные клетки в спинном мозге, что ведет к очередному расслаблению скелет ной мышцы, и т.д. Мышца поочередно со кращается и расслабляется в результате по ступления к ее мотонейронам импульсов от собственных рецепторов. Описанные процес сы в равной степени относятся и к мышце сгибателю, и к мышце-разгибателю, при этом расслабление скелетной мышцы запус кает механизмы ее сокращения, а сокраще ние скелетной мышцы активирует механиз мы, расслабляющие мышцу. Для обеспечения поочередного сгибания и разгибания конечностей при шагательным рефлексе мышцы-сгибатели и разгибатели должны сокращаться и расслабляться после довательно друг за другом, что достигается с помощью торможения центра-aнтaгониста при возбуждении центра-агониста, причем, если на одной ноге сокращены сгибатели, на другой ноге сокращаются разгибатели, что обеспечивается поступлением афферентных импульсов от мышечных и сухожильных ре цепторов и поочередным возбуждением и торможением центров-сгибателей и разгиба телей (рис. 8.3). Шагательные координированн ые дви же ния возможны в отсутствие обратной аффе- | | 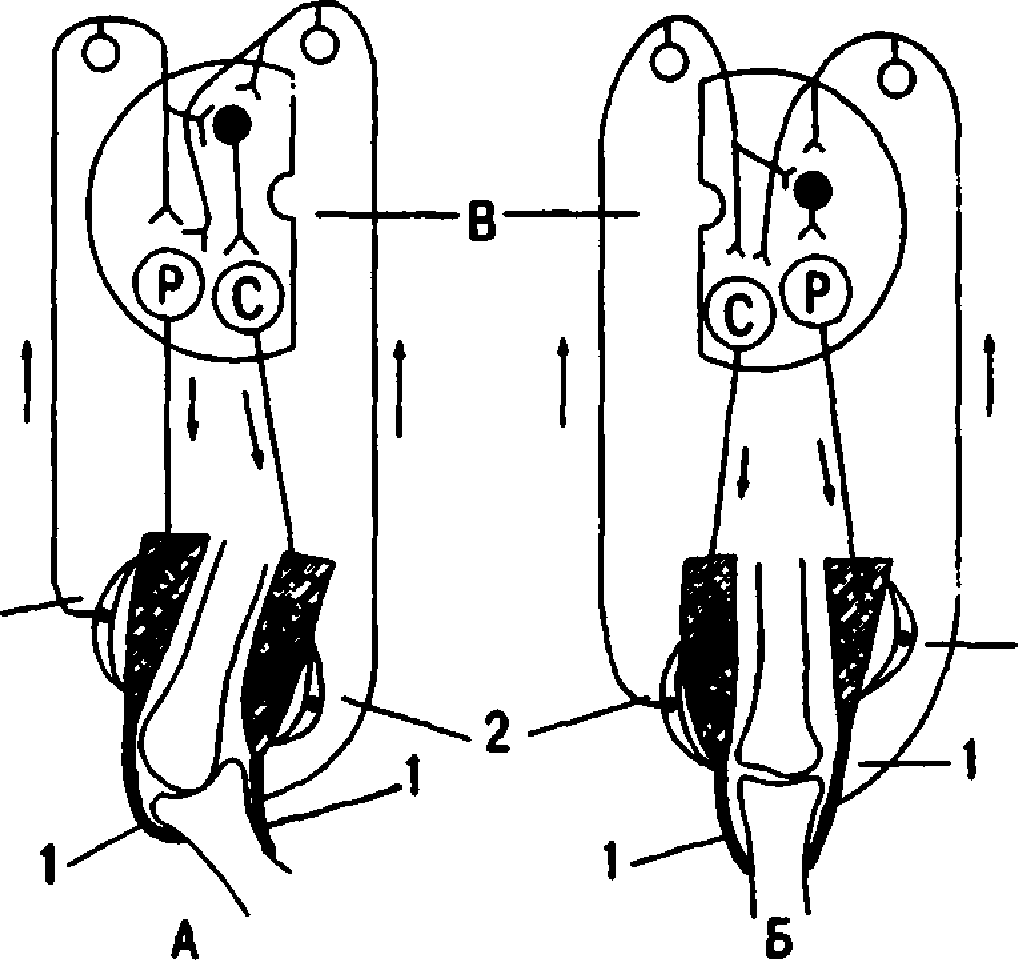 |
| 120 | Рис. 8.3. Рефлекторные дуги шагательного реф лекса (показан коленный сустав). А — конечность согнута; Б — контралатеральнав конеч- ность разогнута; В - полусегмегпы спинного мозга. 1 —сухожилии и их рецепторы (рецепторы Гольджи); 2 - мы шечные рецепторы (мышечныс веретсна); стрелки вверх - афкрснтные пути от проприореиспторов; стрелки вниз -эффсрентныс пути от центров сгибания (С) и раэгибанивi (Р). Светлые — возбуждающие. черные — тормозяшие нейроны. рентации от проприореиепторов. Они осу ществляются с помощью межсегмеЕiтарных связей на уровне спинного мозга. О наличии межсегментарных связей на уровне спинного мозга свидетельствует также факт вовлечения в шагательный рефлекс всех четырех конеч ностей при достаточно длительном и силь ном раздражении одной конечности при ин тактных афферентных путях. 8.1.5. РЕГУЛЯЦИЯ ТОНУСА МЫШЦ Тонус исследуют после поперечной перерез ки спинного мозга. Сразу после перерезки в эксперименте или повреждения его у челове ка в случае травмы наблюдаются мышечная атония и отсутствие рефлексов (спинальный шок). Главной причиной спинального шока является выключение влияния вышележаших отделов ЦНС на спинной мозг (повторная перерезка спинного мозга ниже первой по вторно шок не вызывает). Спинальный шок у лягушек длится несколько минут, у собак - несколько пней, у человека — около 2 мес. После исчезновения спинального шока тонус мышц, иннервируемых посредством сегмен тов спинного мозга, которые находятся ниже перерезки (повреждения), резко повышается. Сгибательные ii разгибательные рефлексы |