Ности. Связывание ангиотензина с рецепторами вызывает увеличение прогiииаемости
 Скачать 458.29 Kb. Скачать 458.29 Kb.
|

|
По-видимому, и Cl— транспортируется в этом —случае из клетки специальной хлорной помпойs, хотя обычно отмечают, что градиент кониент раиии С1— поддерживается отрицательным элект рическим зарядом внутри клетки — С1— вытал кивается отрицательным электрическим заря дом. Поскольку возникновение ТПСП обеспе чивается входом С1— в клетку, это доказывает, что концентрационный градиент действует сильнее противоположно направленного электрического градиента. Отсутствие С1—помпы привело 6ы к накоплению С1— в клетке, наступлению равно весия между электрическими и химическим (кон иентраиионным) градиентами для С1— и нару шению процесса торможения. Но этого не про исходит, что подтверждает наличие хлорной помпы. Как выяснилось, ТПСП могут возникать вследствие уменьшения проницаемости мем браны для ыа*, что также сопровождается ги перполяризацией клеточной мембраны, осо бенно если проницаемость для К' и Crсо храняется прежней. Такого рода ТПСП были зарегистрированы в нейронах симпатических ганглиев. В. Разновидности постсинаптического тор- можения. Обычно выделяют возвратное, ла теральное, параллельное и прямое (реци прокное) постсинаптическое торможение. Имеются и другие варианты классификаций. Некоторые авторы называют только два тор можения -- возвратное и прямое, последнее трактуется по—разному. В реальной действи тельности вариантов торможения больше, они определяются множеством связей раз личных нейронов, в частности их коллатера лей. 1. Возвратное постеинаптическое тормо жение — торможение, при котором тормоз-
108 мозная цепь называется торможением Рен- шоу — в Честь ученого, которым ее открыл,
2. Параллельное торможение может вы полнять подобную же роль, когда возбужде ние блокирует само себя, за счет диверген ции по коллатерали с включением тормозной клетки на своем пути и возвратом импульсов
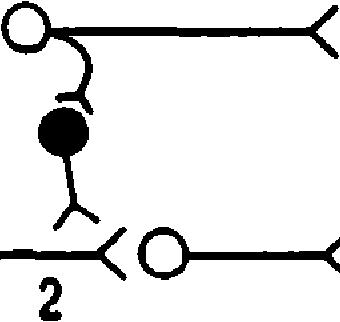
-60 -65 -70'. 0 1 а мс Ра волокон 1а 6 -60 -65 ВПСП -70 0I 1амс Раздрroкеине волокон 1а через 20 мс после АездWажечия воЛОкон 1ь  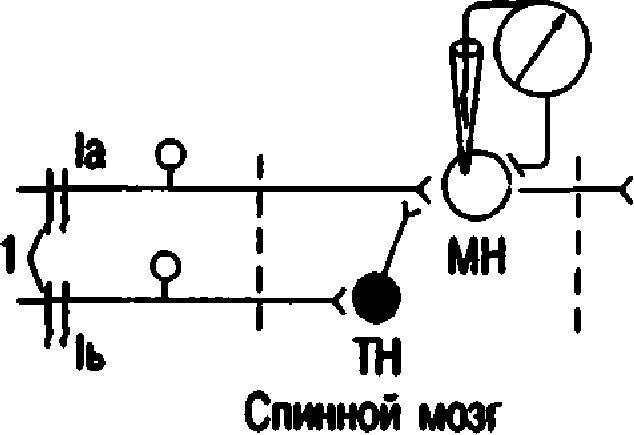 а Рис. 7.9. Опыт, доказываюший на личие пресинаптического торможе ния. А - схема опыта: 1а - ас1хасрентный путь от мышечных рецепторов мыши - разги батгелей (моносинаптическая ре fiлектор ная луга); МН - мотонейрон спинного мозга; 1б - ас ферентный путь от сухо жильных рецепторов мыши - сгибате лей; ТН - тормозящий нсйрон: 1 - раз дражаюшие электроды. G - возбуждение нейрона. В -- низкоаиплнтудный ВПСП, не обеспечивающий возникновения П!1. В 7.8.2. ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ А. Открытие. Пресинаптическое торможение первоначально выявлено также в спинном мозге в опыте с регистрацией активности мо тонейронов моносинаптической рефлектор-
Б. Механизм пресинаптического торможе-
109 яние, естественно, остается неизменным. Де поляризацию пресинаптической терминали вызывают специальные тормозные вставоч ные клетки, аксоны которых образуют си напсы на пресинаптических окончаниях ак сона-мишени (см. рис. 7.9). Торможение (де поляризация) после одного афферентного залпа продолжается 300--400 мс, медиатором является гамма-аминомасляная кислота (ГАМ К), которая действует на ГАМ К,-рецеп- торы. Деполяризация является следствием по- вышения проницаемости для С1-, в результа- те чего он выходит из клетки. По-видимому,
Полагают также, что деполяризация пре синаптических терминалей может возник нуть при накоплении К в межклеточной жидкости в результате повышенной актив ности нервных окончаний и соседних нерв-
В. Разновидности пресинаптического тор можения изучены недостаточно. По-видимо му, имеются те же варианты, что и для пре синаптического торможения. В частности, на
рис. 7.10 представлено параллельное и лате- ральное пресинаптическое торможение. Од- нако возвратного пресинаптического тормо- жения на уровне спинного мозга (по типу возвратного постсинаптического торможе ния) у млекопитающих обнаружить не уда лось, хотя у лягушек оно выявлено. В реальной действительности взаимоотно шения возбуждающих и тормозных нейронов значительно сложнее, чем представлено на рис. 7.8 и 7.10, тем не менее все варианты пре- и постсинаптического торможения можно объединить а две группы: 1) когда блокируется собственный путь самим рас пространяюшимся возбуждением с помощью встааочных тормозных клеток (параллельное и возвратное торможение) и 2) когда блоки руются другие нервные элементы под влия нием импульсов от соседних возбуждающих нейронов с включением тормозных клеток (латеральное и прямое торможение). По скольку тормозные клетки сами могут быть заторможены другими тормозными нейрона ми (торможение), это может облегчить рас пространение возбуждения. 7.8.3. РОЛЬ РАЗЛИЧНЫХ ВИДОВ ТОРМОЖЕНИЯ И ИХ ЛОКАЛИЗАЦИЯ
А. Пре- и постсинаптическое торможение ши роко представлено в различных отделах ЦНС: в частности, глицин — медиатор пост синаптического торможения, кроме клеток Реншоу, обнаружена стволе мозга. ГАМК1 реиепторы локализованы на нейронах гиппо кампа, мозжечка, гипоталамуса, коры боль шого мозга, аксонах первичных афферент-
110 кируется также столбнячным токсином, ко- торый нарушает высвобождение тормозных медиаторов. В заключение необходимо отметить, что в приведенную классификацию видов тормо жения следует внести изменения. Это связа но с тем, что ГАМ К-рецепторы локализую тюся также, кроме пресинаптических терми налей, на нервных волокнах и вне синапсов, на соме и дендритах нейронов, т.е. являются постсинаптическими. В последние годы обнаружены тормозные нейроны, s которых из одного и того же нервного окончания выделяется два медиато ра — ГАМК и глицин. Этот вариант тормоз ных нейронов встречается наиболее часто в спинном мозге и стволе мозга. Таким обра зом, s настоящее время известно три вида тормозных вставочных нейронов: глицинер гические, вызывающие постсинаптическое торможение, ГАМКергические нейроны, вы зывающие пре- и постсинаптическое тормо жение, и тормозные нейроны смешанного типа, выделяющие два медиатора — глииин и ГАМ К. Поэтому классифицировать торможе ние необходимо по двум признакам: по ло кализации (пре- и постсинаптическое) и по природе нейронов (глицинергическое, ГАМ Кергическое и смешанное, табл. 7.1). Т а б л и ц а 7.1. Торможение в ЦНС Реиеп- ' Ионный торы механизм. Блокаторы Вид 1 тормо- Нейроны жения
ческое 1 Постси- ГАМКер- ГАМКI напти- ; гические ГАМК2 ческое Глииинер- Глииин 1 гические Иногда в качестве разновидности центрально го торможения выделяют |