фзл. Общая физиология возбудимых тканей
 Скачать 1.43 Mb. Скачать 1.43 Mb.
|

|
Общая физиология возбудимых тканей Раздражимость и возбудимость. Виды возбудимых тканей и их свойства. Общие и специфические признаки возбуждения. Законы возбуждения (силы, времени и градиента). Классификация раздражителей. Раздражимость – способность реагировать изменением обмена веществ на действие раз-ля Возбудимость – ответ на раз-е возбуждением Возбудимые ткани: нервные мышечные, некоторые секреторные Возбуждение - -ответ на раздражение Специализированные признаки возбуждения: проведение возбуждения нервной тканью, сокращение мышцы, секреция железы Общие признаки возбуждения: Генерация ПД, метаболитические изменения. Раздражители: 1) Естественные (нервные импульсы) 2)Искуственные (физ., хим.,температура ток..) Также раздражители: Адекватные – при минимальных энергозатратах вызывают возбуждение в естественных условиях Неадекватные – вызывают возбуждение при достаточной силе и длительности воздействия Законы возбуждения: 1) Закон силы: Чем больше сила раздражителя, тем сильнее ответная реакция. Однако выраженность ответной реакции растет лишь до определенного максимума. Закону силы подчиняется целостная скелетная, гладкая мышца, так как они состоят из многочисленных мышечных клеток, имеющих различную возбудимость. 2) .Закон силы-длительности.(времени) Между силой и длительностью действия раздражителя имеется определенная взаимосвязь. Чем сильнее раздражитель, тем меньшее время требуется для возникновения ответной реакции. Зависимость между пороговой силой и необходимой длительностью раздражения отражается кривой силы-длительности. По этой кривой можно определить ряд параметров возбудимости. а) Порог раздражения – это минимальная сила раздражителя, при которой возникает возбуждение. б) Реобаза – это минимальная сила раздражителя, вызывающая возбуждение при его действии в течение неограниченно долгого времени. На практике порог и реобаза имеют одинаковый смысл. Чем ниже порог раздражения или меньше реобаза, тем выше возбудимость ткани. в) Полезное время – это минимальное время действия раздражителя силой в одну реобазу, за которое возникает возбуждение. г) Хронаксия – это минимальное время действия раздражителя силой в две реобазы, необходимое для возникновения возбуждения. Этот параметр предложил рассчитывать Л. Лапик, для более точного определения показателя времени на кривой силы-длительности. Чем короче полезное время или хронаксия, тем выше возбудимость и наоборот. В клинической практике реобазу и хронаксию определяют с помощью метода хронаксиметрии для исследования возбудимости нервных стволов. Закон градиента или аккомодации. Реакция ткани на раздражение зависит от его градиента, Т.е. чем быстрее нарастает сила раздражителя во времени тем быстрее возникает ответная реакция. При низкой скорости нарастания силы раздражителя растет порог раздражения. Поэтому если сила раздражителя возрастает очень медленно, возбуждения не будет. Это явление называется аккомодацией. Физиологические особенности мембран возбудимых тканей. Роль мембранных белков. Виды и значение ионных каналов и насосов в функционировании возбудимых клеток. Механизмы деятельности Na+-K+-насоса, Са2+-насоса и Na+-Ca2+-насоса. Ионные каналы и насосы как мишени действия лекарств. Биологические мембраны - функционально активные структуры клеток, ограничивающие цитоплазму и большинство внутриклеточных структур; образуют единую внутриклеточную систему канальцев, складок и замкнутых полостей. Структурная основа мембраны – двойной слой фосфолипидов, в который встроены мембранные белки. Молекулы липидов амфотерны. Своими гидрофильными частями они обращены в сторону водной среды (межклеточная жидкость и цитоплазма), гидрофобные части молекул направлены внутрь липидного бислоя. Белковые молекулы выполняют роль каналов рецепторов, насосов, ферментов. Канал – это белковая макромолекула, образующая пору через двухслойную липидную мембрану. Натриевый канал – тетродотоксин. Калиевый канал – тетраэтиламмоний. Пора - молекулярное динамическое образование. Образована транспортным ферментом, который способен в 200 раз ускорить диффузию. Воротный механизм – на внутренней стороне мембране, это белок, способный к конформации (изменение пространственной конфигурации молекул) Ионные каналы мембраны – Белковые трубочки разного диаметра заполненные жидкостью проходящие мембрану насквозь Сенсор напряжения – белковая молекула в самой мембране, способна реагировать на изменение мембранного потенциала. Селективный фильтр – определяет однонаправленное движение ионов через пору и ее избирательную проницаемость. Пассивный перенос веществ через мембраны проходит без затрат энергии при наличии градиентов (разницы концентраций веществ, разности электрохимического градиента, при наличие градиента давления и осмотического градиента). При этом пассивный транспорт осуществляется с помощью: - диффузии. - фильтрация. Осуществляется при наличии разности гидростатического давления. - осмос. При осмосе происходит движение растворителя. То есть вода из чистого раствора будет переходить в раствор с большей концентрацией. Во всех этих случаях не происходит затраты энергии. Вещества идут через поры, которые имеются в мембране. В мембране существуют поры с медленной проводимостью, но таких пор в мембране не много. Большинство каналов в мембране имеет в своем строении еще воротный механизм, который перекрывает канал. Эти каналы могут управляться двумя способами: реагировать на изменение заряда (электровозбудимые или потенциалозависимые каналы). В другом случае ворота в канале открываются, когда присоединяется химическое вещество (хемовозбудимые или лигандозависимые). Активный перенос веществ через мембрану связан с переносом веществ против градиента. Для активного транспорта используются интегральные белки, которые обладают ферментативными функциями. В качестве энергии используется АТФ. Интегральные белки имеют специальные механизмы (белок), которые активизируется либо при повышении концентрации вещества снаружи клетки, или при понижении внутри. Наиболее важный процесс активного транспорта - это работа Na/K-насоса, существующего практически во всех клетках; насос выкачивает ионы натрия из клетки, одновременно накачивая ионы калия внутрь клетки. Таким образом обеспечивается низкая внутриклеточная концентрация ионов натрия и высокая-калия (табл. 1.1). Градиент концентрации ионов натрия на мембране имеет специфические функции, связанные с передачей информации в виде электрических импульсов, а также с поддержанием других активных транспортных механизмов и регулирования объема клетки (см. ниже). Поэтому неудивительно, что более 1/3 энергии, потребляемой клеткой, расходуется на Na/К-насос, а в некоторых наиболее активных клетках на его работу расходуется до 70% энерги насос—кальциевый; это насос откачивает ионы кальция (Са2+) из клетки и участвует в поддержании их внутриклеточной концентрации на крайне низком уровне (табл. 1.1). Кальциевый насос присутствует с очень высокой плотностью в саркоплазматическом рети-кулуме мышечных клеток, которые накапливают ионы кальция в результате расщепления молекул АТФ. Ионные каналы — порообразующие белки (одиночные либо целые комплексы), поддерживающие разницу потенциалов, которая существует между внешней и внутренней сторонами клеточной мембраны всех живых клеток. Относятся к транспортным белкам. С их помощью ионы перемещаются согласно их электрохимическим градиентам сквозь мембрану. Такие комплексы представляют собой набор идентичных или гомологичных протеинов, плотно упакованных в липидном бислое мембраны вокруг водной поры. Каналы расположены в клеточной и некоторых внутренних мембранах клетки. Через ионные каналы проходят ионы Na+ (натрия), K+ (калия), Cl- (хлора) и Ca++ (кальция). Из-за открывания и закрывания ионных каналов меняется концентрация ионов по разные стороны мембраны и происходит сдвиг мембранного потенциала. Виды ионных каналов: Лиганд-зависимые ионные каналы Эти каналы открываются, когда медиатор, связываясь с их наружними сторонами, меняет их конформацию. Открываясь, они впускают ионы, изменяя этим мембранный потенциал. Лиганд-зависимые каналы почти нечувствительны к изменению мембранного потенциала. Они генерируют электрический сигнал, сила которого зависит от количества медиатора, поступающего в синаптическую щель и времени, которое он там находится. Потенциал-зависимые ионные каналы Эти каналы отвечают за распространение потенциала действия, они открываются и закрываются в ответ на изменение мембранного потенциала. Например, натриевые каналы. Если мембранный потенциал поддерживается на уроне потенциала покоя, натриевые каналы закрыты и натриевый ток отсутствует. Если мембранный потенциал сдвигается в положительную сторону, то натриевые каналы откроются и в клетку начнут входить ионы натрия по градиенту концентрации. Через 0,5 мс после установления нового значения мембранного потенциала, этот натриевый ток достигнет максимума. А еще через несколько миллисекунд падает почти до нуля. Это значит, что каналы через некоторое время закрываются, даже если клеточная мембрана остается деполяризованной. Но закрывшись, они отличаются от состояния, в котором находились до открытия, теперь они не могут открываться в ответ на деполяризацию мембраны то есть они инактивированны. В таком состоянии они останутся до тех пор, пока мембранный потенциал не вернется к исходному значению и не пройдет восстановительный период, занимающий несколько миллисекунд. Важную роль в действии лекарственных веществ играет их способность имитировать или блокировать действие эндогенных лигандов, регулирующих ток ионов через каналы плазматической мембраны. местные анестететики блокируют потенциалозависимые Na -каналы. К числу блокаторов Na -каналов относятся и многие противоаритмические средства. Кроме того, было показано, что ряд противоэпилептических средств (дифенин, карбамазепин) также блокируют потенциалозависимые Na -каналы и с этим связана их противосудорожная активность. Ионы 2+Са принимают участие во многих физиологических процессах: всокращении гладких мышц, в проведении возбуждения по проводящейсистеме сердца, в секреторной активности клеток, в функции2+тромбоцитов и др. Так, верапамил оказывает более сильное влияние на ино-, хронотропную функцию сердца и на атриовентрикулярную проводимость и в меньшей степени на гладкие мышцы сосудов; нифедипин оказывает большее воздействие на гладкие мышцы сосудов и меньшее - на функцию сердца; дилтиазем в равной степени влияет на гладкие мышцы сосудов и проводящую систему; нимодипин обладает избирательным цереброваскулярным действием. Активаторы К -каналов участвуют в механизме их открытия ивыхода ионов К из клетки. Если этот процесс происходит в гладкихмышцах сосудов, то развивается гиперполяризация мембраны, тонус мышц уменьшается и снижается артериальное давление. Такой механизм гипотензивного действия характерен для миноксидила. Природа потенциала покоя (определение и механизмы формирования). Изменение поляризации мембраны при возбуждении и торможении нейронов. потенциалом покоя (ПП) называют разность потенциалов покоящейся клетки между внутренней и наружной сторонами мембраны. Внутренняя сторона мембраны клетки заряжена отрицательно по отношению к наружной. Принимая потенциал наружного раствора за нуль, МПП записывают со знаком «минус». Величина МПП зависит от вида ткани и варьирует от -9 до -100 мв. Следовательно, в состоянии покоя клеточная мембрана поляризована. Уменьшение величины МПП называютдеполяризацией, увеличение — гиперполяризацией, восстановление исходного значения МПП —реполяризацией мембраны. Если на нерв или мышцу нанести раздражение выше порога возбуждения, то МПП нерва или мышцы быстро уменьшится и на короткий промежуток времени (миллисекунда) произойдет перезарядка мембраны: ее внутренняя сторона станет заряженной положительно относительно наружной. Это кратковременное изменение МПП, происходящее при возбуждении клетки, которое на экране осциллографа имеет форму одиночного пика, называется мембранным потенциалом действия (МПД). Поляризация мембраны в покое объясняется наличием открытых калиевых каналов и трансмембранным градиентом концентраций калия, что приводит к выходу части внутриклеточного калия в окружающую клетку среду, т. е. к появлению положительного заряда на наружной поверхности мембраны. Органические анионы – крупномолекулярные соединения, для которых мембрана клетки непроницаема, создают на внутренней поверхности мембраны отрицательный заряд. Поэтому чем больше разница концентраций калия по обе стороны от мембраны, тем больше его выходит и тем выше значения МП. Переход ионов калия и натрия через мембрану по их концентрационному градиенту в конечном итоге должен был бы привести к выравниванию концентрации этих ионов внутри клетки и в окружающей ее среде. Но в живых клетках этого не происходит, так как в клеточной мембране имеются натрий-калиевые насосы, которые обеспечивают выведение из клетки ионов натрия и введение в нее ионов калия, работая с затратой энергии. Они принимают и прямое участие в создании МП, так как за единицу времени ионов натрия выводится из клетки больше, чем вводится калия (в соотношении 3:2), что обеспечивает постоянный ток положительных ионов из клетки. То что выведение натрия зависит от наличия метаболической энергии, доказывается тем, что под действием динитрофенола, который блокирует метаболические процессы, выход натрия снижается примерно в 100 раз. Таким образом, возникновение и поддержание мембранного потенциала обусловлено избирательной проницаемостью мембраны клетки и работой натрий-калиевого насоса. Потенциал действия (ПД) возникает на мембранах возбудимых клеток под влиянием раздражителя пороговой или сверхпороговой величины, который увеличивает проницаемость мембраны для ионов натрия. Ионы натрия начинают входить внутрь клетки, что приводит к уменьшению величины мембранного потенциала – деполяризации мембраны. При уменьшении МП до критического уровня деполяризации открываются потенциалозависимые каналы для натрия и проницаемость мембраны для этих ионов увеличивается в 500 раз (превышая проницаемость для ионов калия в 20 раз). В результате проникновения ионов натрия в цитоплазму и их взаимодействия с анионами разность потенциалов на мембране исчезает, а затем происходит перезарядка клеточной мембраны (инверсия заряда, овершут) – внутренняя поверхность мембраны заряжается положительно по отношению к наружной (на 30 – 50 мВ), после чего закрываются натриевые каналы и открываются потенциалозависимые калиевые каналы. В результате выхода калия из клетки начинается процесс восстановления исходного уровня мембранного потенциала покоя – реполяризация мембраны. Если такое повышение проводимости для калия предотвратить введением тетраэтиламмония, который избирательно блокирует калиевые каналы, мембрана реполяризуется гораздо медленнее. Натриевые каналы можно блокировать тетродотоксином и разблокировать последующим введением фермента проназы, который расщепляет белки. Таким образом, в основе возбуждения (генерации ПД) лежит повышение проводимости мембраны для натрия, вызываемое ее деполяризацией до порогового (критического) уровня. 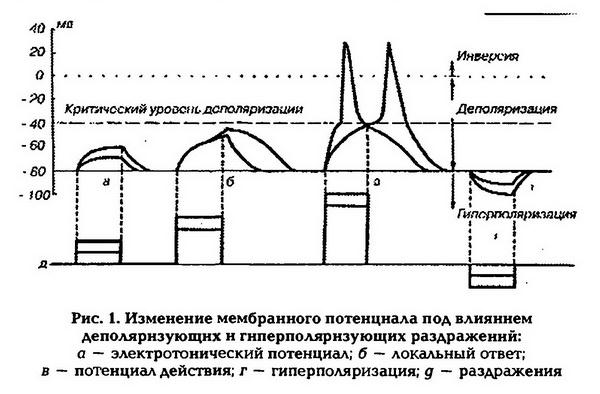 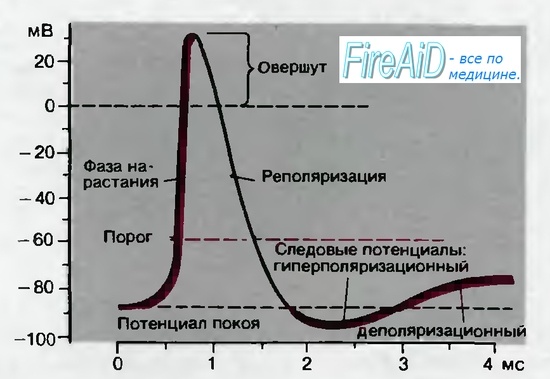 В потенциале действия различают следующие фазы: 1. Предспайк – процесс медленной деполяризации мембраны до критического уровня деполяризации (местное возбуждение, локальный ответ). 2. Пиковый потенциал, или спайк, состоящий из восходящей части (деполяризация мембраны) и нисходящей части (реполяризация мембраны). 3. Отрицательный следовой потенциал – от критического уровня деполяризации до исходного уровня поляризации мембраны (следовая деполяризация). 4. Положительный следовой потенциал – увеличение мембранного потенциала и постепенное возвращение его к исходной величине (следовая гиперполяризация). При увеличении амплитуды подпороговых раздражений от 0,5 до 0,9 пороговой величины развитие деполяризации мембраны происходит не прямолинейно, а по S-образной кривой. Деполяризация продолжает нарастать и после прекращения раздражения, а затем сравнительно медленно исчезает. Этот процесс получил название локального ответа. При увеличении амплитуды подпороговых раздражений от 0,5 до 0,9 пороговой величины развитие деполяризации мембраны происходит не прямолинейно, а по S-образной кривой. Деполяризация продолжает нарастать и после прекращения раздражения, а затем сравнительно медленно исчезает. Этот процесс получил название локального ответа. Локальный ответ имеет следующие свойства: 1. возникает при действии подпороговых раздражителей; 2. находится в градуальной зависимости от силы стимула (не подчиняется закону «все или ничего»); локализуется в месте действия раздражителя и не способен к распространению на большие расстояния; 3. может распространяться лишь локально, при этом его амплитуда быстро уменьшается; 4. локальные ответы способны суммироваться, что приводит к увеличению деполяризации мембраны. В период развития локального ответа возрастает поток ионов натрия в клетку, что повышает ее возбудимость. Локальный ответ является экспериментальным феноменом, однако по перечисленным выше свойствам он близок к таким явлениям, как процесс местного нераспространяющегося возбуждения и возбуждающего постсинаптического потенциала (ВПСП), который возникает под влиянием деполяризующего действия возбуждающих медиаторов. |