фзл. Общая физиология возбудимых тканей
 Скачать 1.43 Mb. Скачать 1.43 Mb.
|

|
Роль сурфактанта Сурфактант противодействует силам поверхностного натяжения альвеол и таким образом поддерживает нужную форму альвеол и облегчает вдох Сурфактант - белково-липидное вещество, вырабатываемое альвеоцитами второго порядка. Выработка увеличивается при действии глюкокартикойдных гормонов. Давление в плевральной щели. Грудная клетка образует герметичную полость, обеспечивающую изоляцию легких от атмосферы. Легкие покрывает висцеральный плевральный листок, а внутреннюю поверхность грудной клетки – париетальная плевра. Между этими листками существует щелевидное пространство, заполненное плевральной жидкостью. Внутриплевральное давление между плевральными листками в норме ниже, чем атмосферное. И это состояние называется отрицательным давлением в плевральной щели (полости). При открытых верхних дыхательных путях давление во всех отделах легких равно атмосферному. Перенос атмосферного воздуха в легкие происходит при появлении разницы давлений между внешней средой и альвеолами легких. При каждом вдохе объем легких увеличивается, давление заключенного в них воздуха или внутрилегочное давление становится ниже атмосферного на 6-9 мм рт. ст. и воздух засасывается в легкие. При выдохе уменьшается объем легких, давление в альвеолах становится выше атмосферного и альвеолярный воздух выходит во внешнюю среду. На высоте спокойного выдоха отрицательное давление в плевральной щели составляет 1,5-3 мм рт. ст. Отрицательное давление в плевральной полости обусловлено так называемой эластической тягой легких –силой, с которой легкие постоянно стремятся уменьшить свой объем. Аэродинамическое сопротивление обусловлено наличием силы трения между молекулами газовой смеси и поверхностью дыхательных путей. Аэродинамическое сопротивление увеличивается при обструктивных поражениях дыхательной системы (отек слизистой бронхов, бронхоспазм, хронические воспалительные заболевания легких и др.). Частным случаем аэродинамического сопротивления является сопротивление, не связанное непосредственно с системой органов дыхания (приложенное извне), например, сопротивление интубационной трубки или трахеотомической канюли. Вопрос 46. Газообмен в легких. Газообмен в легких совершается между альвеолярным воздухом и кровью путем диффузии. Альвеолы легких оплетены густой сетью капилляров. Стенки альвеол и капилляров очень тонкие, что способствует проникновению газов из легких в кровь и наоборот. Газообмен зависит от величины поверхности, через которую осуществляется диффузия газов, и разности парциального давления (напряжения) диффундирующих газов. При глубоком вдохе альвеолы растягиваются, и их поверхность достигает 100-105 м2. Так же велика и поверхность капилляров в легких. Есть, и достаточная, разница между парциальным давлением газов в альвеолярном воздухе и напряжением этих газов в венозной крови. Факторы: Уровень вентиляции в легких Перфузия легких (кровоснабжение) Особенности диффузии газов через аэрогематический барьер (определяется разницей парциальный давлений газов, и составляет аэрогематический барьер) Парциальное давление газов – давление, под которым газ находится в газовой среде. Если характеризуется содержание газов в жидкости используется понятие парциальное напряжение. Особенности газообмена в тканях. Обмен газов в тканях осуществляется в капиллярах. Через их тонкие стенки кислород поступает из крови в тканевую жидкость и затем в клетки, а углекислота из тканей переходит в кровь. Концентрация кислорода в крови больше, чем в клетках, поэтому он легко диффундирует в них. Концентрация углекислого газа в тканях, где он собирается, выше, чем в крови. Поэтому он переходит в кровь, где связывается химическими соединениями плазмы и отчасти с гемоглобином, транспортируется кровью в легкие и выделяется в атмосферу. Вопрос 47. Транспорт газов кровью. Кровь переносит кислород и углекислый газ. В крови, как и во всякой жидкости, газы могут находиться в двух состояниях: в физически растворенном и химически связанном. И кислород и углекислый газ в очень небольшом количестве растворяются в плазме крови. Большая часть кислорода и углекислого газа переносится в химически связанном виде. Основной переносчик кислорода - гемоглобин крови. 1 г гемоглобина связывает 1,34 мл кислорода. Гемоглобин обладает способностью вступать в соединение с кислородом, образуя оксигемоглобин. Чем выше парциальное давление кислорода, тем больше образуется оксигемоглобина. В альвеолярном воздухе парциальное давление кислорода 100-110 мм рт. ст. При таких условиях 97% гемоглобина крови связывается с кислородом. Кровь приносит к тканям кислород в виде оксигемоглобина. Здесь парциальное давление кислорода низкое, и оксигемоглобин - соединение непрочное - высвобождает кислород, который используется тканями. На связывание кислорода гемоглобином оказывает влияние и напряжение углекислого газа. Углекислый газ уменьшает способность гемоглобина связывать кислород и способствует диссоциации оксигемоглобина. Повышение температуры также уменьшает возможности связывания гемоглобином кислорода. Известно, что температура в тканях выше, чем в легких. Все эти условия помогают диссоциации оксигемоглобина, в результате чего кровь отдает высвободившийся из химического соединения кислород в тканевую жидкость. При специальной тренировке организм может приспосабливаться к пониженному содержанию кислорода в атмосферном воздухе. У тренированного человека углубляется дыхание, увеличивается количество эритроцитов в крови за счет усиленного образования их в кроветворных органах и поступления из депо крови. Кроме того, усиливаются сердечные сокращения, что приводит к увеличению минутного объема крови. Для тренировки широко применяют барокамеры. Углекислый газ переносится кровью в виде химических соединений - бикарбонатов натрия и калия. Связывание углекислого газа и отдача его кровью зависят от его напряжения в тканях и крови. Кроме того, в переносе углекислого газа участвует гемоглобин крови. В капиллярах тканей гемоглобин вступает в химическое соединение с углекислым газом. В легких это соединение распадается с освобождением углекислого газа. Около 25-30% выделяемого в легких углекислого газа переносит гемоглобин. Кривая диссоциации оксигемоглобина 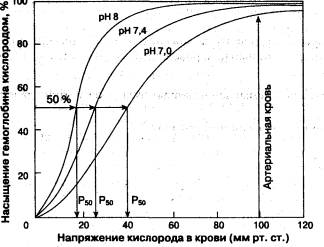 Сродство гемоглобина к кислороду Эффект рН и температуры на сродство гемоглобина к кислороду. Молекулы гемоглобина способны реагировать с ионами водорода, в результате этой реакции происходит снижение сродства гемоглобина к кислороду. При насыщении гемоглобина менее 100 % низкое рН понижает связывание кислорода с гемоглобином — кривая диссоциации оксигемоглобина смещается вправо по оси х. Это изменение свойства гемоглобина под влиянием ионов водорода называется эффектом Бора. Метаболически активные ткани продуцируют кислоты, такую как молочная, и С02. Если рН плазмы крови снижается от 7,4 в норме до 7,2, что имеет место при сокращении мыщц, то концентрация кислорода в ней будет возрастать вследствие эффекта Бора. Например, при постоянном рН 7,4 кровь отдавала бы порядка 45 % кислорода, т. е. насыщение гемоглобина кислородом снижалось до 55 %. Однако когда рН снижается до 7,2, кривая диссоциации смещается по оси х вправо. В результате насыщение гемоглобина кислородом падает до 40 %, т. е. кровь может отдавать в тканях до 60 % кислорода, что на 1/з больше, чем при постоянном рН. Метаболически активные ткани повышают продукцию тепла. Повышение температуры тканей при физической работе изменяет соотношение фракций гемоглобина в эритроцитах и вызывает смещение кривой диссоциации оксигемоглобина вправо вдоль оси х. В результате большее количество кислорода будет освобождаться из гемоглобина эритроцитов и поступать в ткани. Эффект 2,3-дифосфоглицерата (2,3-ДФГ) на сродство гемоглобина к кислороду. При некоторых физиологических состояниях, например при понижении Р02 в крови ниже нормы (гипоксия) в результате пребывания человека на большой высоте над уровнем моря, снабжение тканей кислородом становится недостаточным. При гипоксии может понижаться сродство гемоглобина к кислороду вследствие увеличения содержания в эритроцитах 2,3-ДФГ. В отличие от эффекта Бора, уменьшение сродства гемоглобина к кислороду под влиянием 2,3-ДФГ не является обратимым в капиллярах легких. Однако при движении крови через капилляры легких эффект 2,3-ДФГ на снижение образования оксигемоглобина в эритроцитах (плоская часть кривой диссоциации оксигемоглобина) выражен в меньшей степени, чем отдача кислорода под влиянием 2,3-ДФГ в тканях (наклонная часть кривой), что обусловливает нормальное кислородное снабжение тканей. КИСЛОРОДНАЯ ЕМКОСТЬ КРОВИ - максимальное количествокислорода, обратимо связанное кровью; выражается в объемных процентах; зависит от концентрации в кровигемоглобина. Кислородная емкость крови человека ок. 18-20%. Оксигемометрия— фотометрический метод измерения степени насыщения артериальной крови кислородом, т. е. процентного содержания в ней гемоглобина в форме оксигемоглобина. Оксигемометрия позволяет судить, насколько полно осуществляется оксигенация крови в легких, так как она дает объективный показатель эффективности внешнего дыхания и кровообращения. Оксигемометрию используют для выявления гипоксемической циркуляторной гипоксии (см.). О. производят специальными приборами — оксигемометрами. Пульсоксиметрия — методика определения количества кислорода, связанного с гемоглобином, в артериальной крови. К каждой молекуле гемоглобина может присоединится до четырех молекул кислорода. Средний процент насыщения молекул гемоглобина является кислородной сатурацией крови. 100% сатурация означает, что ккаждая молекула гемоглобина в исследуемом объеме крови переносит четыре молекулы кислорода. Вопрос 48. 1885г. Миславский Н.А. «Д.Ц. – центр жизни.» Д.Ц. – парное образование расположенное в продолговатом мозге справа и слева от его дорсальной срединной борозды и обеспечивающее автоматическую смену фаз дыхания. Виды нейронов дыхательного центра H-пейсмекеры – автоматические нейроны. Инспираторные нейроны (И.Н.) Экспираторные нейроны (Э.Н.) Переходные (инспираторно-экспираторные нейроны, И.-Э., Э.-И.) Нейроны дыхательного центра образуют скопление в виде дыхательных или респираторных функций. Дорсальная респираторная группа. Имеет медиальное расположение В основном нейроны вдоха (И.Н.) Контролирует мотонейроны диафрагмы Обеспечивает рефлекс Геринга-Брейера (рефлекс переключения вдоха на выдох) Получает сигналы от рецепторов растяжения легких по блуждающим нервам Получает сигналы от рецепротов растяжения легких по блуждающим нервам Получает сигналы от верхних структур Г.М., а так же от хемо- и баро- рецепторов сосудов. Вентральная респираторная группа VRG ИН+ЭК Передает сигналы к мотонейронам для наружных и внутренних межреберных мышц 3-pre-Botzinger – комплекс (пейсмекеры) Botzinger-complex Э.Н. Содержит пейсмекерные нейроны  Теории дыхательногоритмогенеза Вопрос 49. Механизмы регуляции дыхания. Центральный (рефлекторный) Наиболее важные рецепторы: А) R-рястяжения (запуск инспираторно-тормозный рефлекс) Б) Ирритантные (калиевые) расположены в стенках верхних и нижних дыхательных путей быстроадаптирующиеся. Возбуждающиеся на действие посторонних раздражителей: пыль, табачный дым и другие. Обеспечивает защитные дыхательные рефлексы: кашель, чихание, а так же бронхоспазм, частое и поверхностное дыхание при повышенной продолжительности вдоха. В) Юксатаальвеолярные (J-R) Практически не адаптируются. Возбуждаются при патологии легких (отек, пневмония, повышенное давление в малом круге, легочная эмболия.) Вызывает бронхоспазм, тахипное, оттдышка. Эти рецепторы чувствительны к гистамину, никотину, простагландинам. Г) Проприорецепторы скелетных мышц Активируются при мышечных нагрузках, вызывают гипервентиляцию легких. Д) Хеморецепторы (улавливают количество газов) Периферические (за пределами ЦНС) в особых зонах сосудистой системе и наиболее чувствительны к снижению PO2 в крови Центральные (В WYC) Представляет собой нейроны, которые активируются при повышении PCO2 и Н+ в межклеточной жидкости мозга Гуморальный. Глу-, Т3, Т4 (тереойдные гормоны), кортизол, аспартат Оказывает возбуждающее действие на дыхательный центр ГАМК-, глицин, опиойды – угнетают деятельность Д.Ц. Высшая форма регуляции дыхания произвольная, обеспечивается корой мозга. Впервые особую роль коры описал Сеченов в 1950г. Он показал приспособительное значение коры мозга для дыхания. Вопрос 50. Функции сердца. Насосная – движение крови по сосудам У.Гарвей впервые отметил эту функцию (1826г.) и открыл круги кровообращения Секреторная (предсердный натрийретическийпептид – облегчает работу сердца: повышает АД, снижает ОЦК, снижает Na+, снижает РААС, снижает гипертрофию миокарда) Поддержание pHкрови – активное потребление лактата при мышечной нагрузке. Предохраняет организм от ацидоза Резервуарная: Конечно-диастолический объем (КДО) объем крови в желудочке к концу диастолы (около 150мл) Конечно-диастолический (КСО) объем крови к концу систолы (около 80мл) Основные показатели деятельности сердца. Ритм ЧСС 70-80 уларов в минуту СисталическийV(CO) или ударный (УО) около 70мл. Тот Vкрови который камера выбрасывает в сосуд CO= КДО-КСО МинутныйVкровообращения – количество крови, проходит через сердце за 1 минуту. МОК= ЧССxУО= 4-6 литров в минуту Сердечный индекс (СИ) = МОК/S = 2-4литра в минуту Тот объем общего кровотока, который за единицу времени приходится на единицу площади Фракция выброса (ФВ) ФВ= УО/KДО – не менее 45% Свойства миокарда. Автоматия Возбудимость Проводимость Сократимость Вопрос 51. Проводящая система сердца образована двумя видами специализированных клеток. Один вид клеток (Р-клетки) обладает автоматизмом, т.е. способностью спонтанно вырабатывать электрические импульсы. Частота возникновения ипульсов зависит от места расположения Р-клеток - чем ближе находятся Р-клетки к началу проводящей системы, тем чаще возникают в них импульсы и, наоборот, чем дальше находятся Р-клетки от начала проводящей системы, тем реже могут возникать в них электрические импульсы. Второй вид (Т-клетки) обладает проводимостью, т.е. способностью проведения возникающих импульсов к сократительному миокарду. Проводящая система сердца начинается синусовым узлом, который расположен в верхней части правого предсердия. Его длина 10-20 мм, ширина 3-5 мм. Именно в нем возникают импульсы, которые вызывают возбуждение и сокращение всего сердца. Нормальный автоматизм синусового узла составляет 50-80 импульсов в минуту. Синусовый узел является автоматическим центром I порядка. Импульс, возникший в синусовом узле мгновенно распространяется по предсердиям, заставляя их сократиться. Но распространиться дальше и сразу же возбудить желудочки сердца эта волна не может, так как миокард предсердий и желудочков разделен фиброзной тканью, которая не пропускает электрические импульсы. И только в одном месте этой преграды не существует. Туда и устремляется волна возбуждения. Но именно в этом месте находится следующий узел проводящей системы, который называется атриовентрикулярным (длина около 5 мм, толщина - 2 мм). В нем происходит задержка волны возбуждения и фильтрация входящих импульсов. Далее нижняя часть узла, утончаясь, переходит в пучок Гиса (длина 20 мм). В последующем пучок Гиса разделяется на две ножки - правую и левую. Правая ножка проходит по правой стороне межжелудочковой перегородки и разветвляясь ее волокна (волокна Пуркинье) пронзают миокард правого желудочка. Левая ножка проходит по левой половине межжелудочковой перегородки и делится на переднюю и заднюю ветви, которые снабжают волокнами Пуркинье миокард левого желудочка. После задержки в результате прохождения атриовентрикулярного узла волна возбуждения, распространяясь по ножкам пучка Гиса и волокнам Пуркинье, мгновенно охватывает всю толщу миокарда желудочков, вызывая их сокращение. Задержка импульса имеет огромное значение и не дает сократиться предсердиям и желудочкам одновременно - сперва сокращаются предсердия, и только вслед за этим - желудочки сердца. В атриовентрикулярном узле, так же как и в синусовом узле, имеются два вида клеток - Р и Т. Атриовентрикулярный узел вместе с начальной частью пучка Гиса является автоматическим центром II порядка, который может самостоятельно вырабатывать импульсы с частотой 35-50 в минуту. Конечная часть пучка Гиса, его ножки и волокна Пуркинье также обладают автоматизмом, однако могут вырабатывать импульсы лишь с частотой 15-35 в минуту и являются автоматическим центром III порядка. Между автоматическими центрами I, II и III порядков возникают следующие взаимодействия. В норме импульс, возникающий в синусовом узле, распространяется на предсердия и желудочки, вызывая их сокращения. Проходя на своем пути автоматические центры II и III порядков импульс каждый раз вызывает разрядку этих центров. После этого в автоматических центрах II и III порядков снова начинается подготовка очередного импульса, которая каждый раз вновь прерывается после прохождения возбуждения из синусового узла. По сути дела, в норме автоматический центр I порядка подавляет активность автоматических узлов II и III порядков. И только в случае отказа синусового узла или нарушения проведения его импульсов на нижележащие отделы включается автоматический узел II порядка, а при его отказе - автоматический узел III порядка. |