фзл. Общая физиология возбудимых тканей
 Скачать 1.43 Mb. Скачать 1.43 Mb.
|

|
Природа потенциала действия, характеристика его фаз. Закон «Всё или ничего». Изменение возбудимости в процессе возбуждения. Механизм кодирование информации в нервной системе. При возбуждении клетки на ее мембране генерируются кратковременные изменения мембранного потенциала, которые называют потенциалом действия (ПД]. фазы ПД . стадия покоя. Эта стадия представлена мембранным потенциалом покоя, который предшествует потенциалу действия. Мембрана во время этой стадии поляризована в связи с наличием отрицательного мембранного потенциала, равного -90 мВ. Фаза деполяризации. В это время мембрана внезапно становится высокопроницаемой для ионов натрия, позволяя огромному числу положительно заряженных ионов натрия диффундировать внутрь аксона. Нормальное поляризованное состояние в -90 мВ нейтрализуется поступающими внутрь положительно заряженными ионами натрия, в результате потенциал стремительно нарастает в положительном направлении. — продолжается 0,2-0,5 мс. Фаза реполяризации. В течение нескольких долей миллисекунды после резкого повышения проницаемости мембраны для ионов натрия, натриевые каналы начинают закрываться, а калиевые — открываться. В результате быстрая диффузия ионов калия наружу восстанавливает нормальный отрицательный мембранный потенциал покоя. фаза следовых потенциалов включает: фазу следовой деполяризации и фазу следовой гиперполяризации. Гиперполяризация — это увеличение разницы мембранного потенциала между наружной и внутренней сторонами мембраны по сравнению с потенциалом покоя, деполяризация — уменьшение мембранного потенциала. Эта фаза в целом длится 2-3 мс. 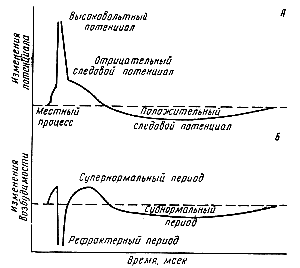 Длительность потенциала действия составляет: в нервах около 1 мс; в нейронах — 2-4 мс; в скелетных мышцах — около 10-20 мс; в кардиомиоците — 200-300 мс Длительность потенциала действия составляет: в нервах около 1 мс; в нейронах — 2-4 мс; в скелетных мышцах — около 10-20 мс; в кардиомиоците — 200-300 мсПериоду возникновения и развития пика потенциала действия т полное исчезновение возбудимости, получившее название абсолютной рефрактерности. В эту фазу второе раздражение не способно вызвать новый потенциал действия, как бы сильно ни было это раздражение. Длительность абсолютной рефрактерности варьируется. Вторая фаза относительной рефрактерности. В это время возбудимость постепенно возвращается к первоначальному уровню, имевшемуся до начала первого раздражения. В период относительной рефрактерности нервное или мышечное волокно способно ответить на сильное раздражение, но амплитуда потенциала действия оказывается резко сниженной. В нервной системе сенсорная информация кодируется путем неимпульсного и импульсного кодирования. Неимпульсное кодирование основано на изменении рецепторного, синаптического или мембранного потенциалов. В его основе лежат процессы, связанные с чувствительностью рецепторов к раздражителю, интенсивностью р-ля, явлениями суммации. (Доминирующий видом в НС) импульсное кодирование, которое осуществляется путем частотного и интервального кодирования. Частотное кодирование происходит за счет количества импульсов в единицу времени, количеством импульсов, следующих последовательно в «пачке», а интервальное — различными временными интервала¬ми между импульсами Механизмы проведения возбуждения по мякотным и безмякотным нервным волокнам. Законы проведения возбуждения. Физиология парабиоза. Физиологические механизмы местной анестезии. При нанесении раздражения возникает деполяризация в области ближайшего перехвата Ранвье - А. Соседний перехват Ранвье - Б находится в состоянии поляризации. Между перехватами возникает разность потенциалов, которая приводит к появлению круговых токов Ионный поток в аксоплазме и в окружающей среде течет от плюса к минусу. Выход круговых токов в перехвате Б приводит к его деполяризации и возникновению потенциала действия. Далее за счет круговых токов возбуждаются последующие перехваты Ранвье. возбуждение в мякотных нервных волокнах передается скачкообразно (сальтаторно) от одного перехвата Ранвье к другому, Сальтаторный способ передачи возбуждения более экономичен, нежели распространение возбуждения по безмякотным нервным волокнам. Возбуждение по мякотным нервным волокнам распространяется без затухания. Скорость распространения возбуждения по мякотным нервным волокнам гораздо выше, чем по безмякотным. Так, скорость распространения возбуждения по двигательным нервным волокнам (мякотные нервы) составляет 80-120 м/с, по волокнам, не покрытым миелиновой оболочкой, - от 0,5 до 2 м/с. В безмякотном волокне возбуждение распространяется медленно, потенциалы действия небольшие, хотя оболочка волокна тонкая, импульсы все равно передаются изолированно. Проведение возбуждения по нервам подчиняется следующим законам: 1.Закон анатомической и физиологической целостности нерва. Первая нарушается при перерезке, вторая – действии веществ, блокирующих проведение, например новокаина. 2.Закон двустороннего проведения возбуждения. Оно распространяется в обе стороны от места раздражения. В организме чаще всего возбуждение по афферентным путям оно идет к нейрону, а по эфферентным – от нейрона. Такое распространение называется ортодромным. Очень редко возникает обратное, или антидромное, распространение возбуждения. 3.Закон изолированного проведения. Возбуждение не передается с одного нервного волокна на другое, входящее в состав этого же нервного ствола. 4.Закон бездекрементного проведения. Возбуждение проводится по нервам без декремента, т.е. затухания. Следовательно, нервные импульсы не ослабляются проходя по ним. Парабиоз (в пер.: “para” - около, “bio” - жизнь) – это состояние на грани жизни и гибли ткани, возникающее при воздействии на нее токсических веществ таких как наркотиков, фенола, формалина, различных спиртов, щелочей и других, а также длительного действия электрического тока. Учение о парабиозе связано с выяснением механизмов торможения, которое лежит в основе жизнедеятельности организма. Фазы. 1. Фаза кратковременного повышения возбудимости. Редко улавливается и заключается в том, что под действием подпорогового раздражителя мышца сокращается. 2. Фаза уравнительная (трансформации). Проявляется в том, что на частые и редкие стимулы мышца отвечает одинаковым по величине сокращением. Выравнивание силы мышечных эффектов происходит, по данным Введенского, за счет парабиотического участка, в котором снижается лабильность под влиянием KСl. более частые сигналы задерживаются в парабиотическом участке, т. к. часть из них попадает в период рефрактерности, который создается предыдущим импульсом и в связи с этим не проявляет своего действия. 3. Парадоксальная фаза. при действии частых стимулов наблюдается слабый сократительный эффект мышцы или вообще его не наблюдается. В то же самое время, на действия редких импульсов имеет место несколько большее по величине сокращение мышцы, чем на более частые. Парадоксальная реакция мышцы связана с еще большим уменьшением лабильности в парабиотическом участке, который практически теряет свойство проводить частые импульсы. 4. Тормозная фаза. В этот период состояния ткани через парабиотический участок не проходят ни частые, ни редкие импульсы, в результате чего мышца не сокращается. Если прекратить действовать KСl, то нервно-мышечный препарат постепенно восстанавливает свою функцию, проходя стадии парабиоза в обратном порядке, или действовать на него одиночными электрическими стимулами, на которые мышца слегка сокращается. К местным анестетикам относятся лекарственные средства, которые в определенных концентрациях блокируют нервную проводимость. Местные анестетики предупреждают генерацию и проведение нервных импульсов. Их основная точка приложения — мембрана нервных клеток. Механизм действия местных анестетиков выражается в блокировке нервного проведения путем вмешательства в процессы генерации нервного потенциала действия. Они уменьшают проницаемость клеточной мембраны для ионов натрия, что сопровождается деполяризацией мембраны. Одним из важнейших путей блокирования натриевых каналов является вытеснение местными анестетиками Са2+ из рецепторов, расположенных на внутренней поверхности мембраны. Изменяя потенциал действия в мембранах нервных клеток, местные анестетики не приводят к выраженному изменению потенциала покоя. Местные анестетики обычно оказывают воздействие только в месте введения. Для снижения системного действия, токсического эффекта и пролонгирования эффекта местных анестетиков их часто используют в комбинации с вазоконстрикторами (адреналин). Местные анестетики делятся на два типа: эстеры (новокаин), метаболизирующиеся в плазме эстеразами, и амиды (лидокаин) Особенности структурно-функциональной организации электрического синапса (эфапса). Механизм передачи информации в электрическом синапсе. Механизм проведения возбуждения аналогичен механизму проведения возбуждения в нервном волокне. Во время развития ПД происходит реверсия заряда пресинаптической мембраны. Электрический ток, возникающий между пресинаптической и постсинаптической мембраной, раздражает постсинаптическую мембрану и вызывает генерацию в ней ПД Особенности ультраструктуры электрического синапса: •узкая (около 5 нм) синаптическая щель; •наличие поперечных канальцев, соединяющих пресинаптическую и постсинаптическую мембрану. Электрические синапсы имеют щель, которая на порядок меньше, чем щель у химических синапсов. Они проводят сигнал в обе стороны без синаптической задержки. Передача сигнала не блокируется при удалении ионов Са2+. Кроме того, электрические синапсы малочувствительны к фармакологическим препаратам и ядам, практически неутомляемы, как и нервное волокно. Контактирующие мембраны нейронов связаны друг с другом полуканалами белковой природы - коннексонами (от англ, соппеаюп - связь). Через коннексоны клетки обмениваются некоторыми компонентами цитоплазмы: аминокислотами, пептидами, РНК, метаболитами, циклическими нуклеотидами. Очень низкое удельное сопротивление сближенных пре- и постсинаптических мембран обеспечивает Хорошую электрическую проводимость. У электрического синапса клеточные мембраны соседних нейронов тесно прилегают друг к другу, образуя узкую щель шириной 2 им. Зона этого сближения мембран называется щелевым контактом. -коннексон- специфические белковые комплексы, состоящие из шести субъединиц и располагающиеся в таком порядке, что в их центре образуется пора, заполненная водой, которая проходит через бислой клеточной мембраны. Через электрический синапс сигнал передается электрическим путем посредством возникновения локальных токов между двумя клетками. Структура и физиологические свойства химического синапса. Механизм передачи информации в химическом синапсе. Химический синапс как «мишень» действия лекарственных средств. Понятие об ионотропных и метаботропных рецепторах. Особенности ультраструктуры химического синапса: • широкая (20–50 нм) синаптическая щель; • наличие в синаптической бляшке синаптических пузырьков (везикул), заполненных химическим веществом, при помощи которого передается возбуждение; • в постсинаптической мембране имеются многочисленные хемочувствительные каналы (в возбуждающем синапсе – для Nа+ , в тормозном – для Cl – и К +), но отсутствуют потенциалчувствительные каналы. На пресинаптической мембране осуществляется трансформация электрического сигнала в химический, который на постсинаптической мембране снова трансформируется в электрический сигнал. Синтез медиатора. Медиатором (посредником) называют химическое вещество, которое обеспечивает одностороннюю передачу возбуждения в химическом синапсе. Некоторые медиаторы (например, ацетилхолин) синтезируются в цитоплазме синаптического окончания, и там же молекулы медиатора депонируются в синаптических пузырьках. Другие медиаторы синтезируются и упаковываются в везикулы в теле нейрона, готовые синаптические пузырьки доставляются в синаптичекую бляшку за счет быстрого аксонного транспорта. Секреция медиатора. Содержимое синаптических пузырьков может выбрасываться в синаптическую щель путем экзоцитоза. При опорожнении одного синаптического пузырька в синаптичекую щель выбрасывается порция (квант) медиатора. Для активации экзоцитоза необходимы ионы Са+2 . В состоянии покоя уровень Са+2 в синаптическом. Приход в синаптическое окончание возбуждения приводит к деполяризации пресинаптической мембраны и открытию потенциал чувствительных Са+2 -каналов. Ионы Са++ поступают в цитоплазму синаптического окончания и активируют опорожнение синаптических пузырьков в синаптическую щель Взаимодействие медиатора с рецепторами постсинаптической мембраны. Молекулы медиатора диффундируют через синаптическую щель и достигают постсинаптической мембраны, где связываются с рецепторами хемочувствительных Na+ -канало. Присоединение медиатора к рецептору приводит к открытию Na+ -каналов, через которые в клетку входят ионы Na+. В результате входа в клетку положительно заряженных ионов происходит локальная деполяризация постсинаптической мембраны, которую называют возбуждающий постсинаптический потенциал (ВПСП) Инактивация медиатора. Ферменты, находящиеся в синаптической щели, разрушают молекулы медиатора. В результате происходит закрытие Na+ -каналов и восстановление МП постсинаптической клетки. Некоторые медиаторы (например, адреналин) не разрушаются ферментами, а удаляются из синаптической щели путем быстрого обратного всасывания (пиноцитоза) в синаптическое окончание. Генерация ПД. В нейро-мышечном синапсе амплитуда единичного ВПСП достаточно велика. Поэтому для генерации ПД в мышечной клетке достаточно прихода одного нервного импульса. Генерация ПД в мышечной клетке происходит в области, окружающей постсинаптическую мембрану. Проведение возбуждения в химическом синапсе одностороннее. Высокая утомляемость. Низкая лабильность. Длинная синаптическая задержка. Чувствительность к действию хим. Агентов. В хим. Синапсе трансформируется ритм ПД. Средства, влияющие на м-холинорецепторы М-холиномиметики (мускариномиметические средства) Пилокарпина гидрохлорид Ацеклидин М-холиноблокаторы (антихолинергические, атропиноподобные средства) Атропина сульфат Метацин Действие большинства антидепрессантов связано с их влиянием на обмен медиаторов-моноаминов, особенно норадреналина и серотонина, и на рецепторы этих медиаторов Во многом он связан с тем, что у ингибиторов обратного захвата серотонина — мощных антидепрессантов — было обнаружено выраженное анксиолитическое действие. Эти препараты помогают при паническом расстройстве, социофобии и других фобиях, неврозе тревоги и неврозе навязчивых состояний, а также при импульсивных расстройствах, по-видимому, близких к данной группе. При тревожных расстройствах используются также бензодиазепины, азапироны и другие транквилизаторы. Ионотропные рецепторы.Вследствие образования постсинаптического потенциала происходит открытие соответствующего ионного канала или сразу при действии медиатора, или через активацию G-белка. При этом рецептор или сам образует ионный канал, или связан с ним. После присоединения лиганда и активации рецептора происходит открытие канала для соответствующего иона. В результате на мембране образуется постсинаптический потенциал. Ионотропные рецепторы – это путь быстрой передачи сигнала и образования ПСП без изменения процессов метаболизма в клетке. Метаботропные рецепторы. Это более сложный путь передачи сигнала. При этом после связывания лиганда с рецептором происходит активация каскада фосфорилирование-дефосфорилирование. Это осуществляется или прямо, или через вторичные посредники, например, через тирозинкиназу, или через цАМФ, или цГМФ, или за счет увеличения внутриклеточного кальция, что в результате приводит к активации протеинкиназ. Фосфорилирование чаще всего включает в себя активацию цАМФ-зависимой или диацилглицерол-зависимой протеинкиназы. Эти эффекты развиваются более медленно и длятся более долго. Сродство рецептора к соответствующему нейромедиатору может меняться так же, как и к гормонам, например, за счет аллостерических изменений рецептора или других механизмов. Поэтому сейчас рецепторы обозначают как мобильные и легко изменяемые структуры. Входя в состав мембраны, белки-рецепторы могут взаимодействовать с другими мембранными белками. Нейромодуляторы, как и нейромедиаторы, могут влиять на число и чувствительность рецепторов. Длительное присутствие больших количеств нейромедиатора или нейромодулятора может снижать их чувствительнос, а недостаток лигандов повышать их чувствительность (ап-регуляция). Нейротрансмиттеры и их классификация. Особенности структурно-функциональной организации рецепторов к нейротрансмиттерам на примере NMDA-рецептора к глутамату и ГАМКА-рецептора к гамма-аминомасляной кислоте. Понятие об агонистах и антагонистах. Рецепторы к нейромедиаторам располагаются на мембранах нейронов или клеток-мишеней (мышечные или железистые клетки). На пресинаптических мембранах чаще располагаются так называемые ауторецепторы, которые регулируют выделение этого же медиатора из пресинаптического окончания. Но есть и гетероауторецепторы, которые регулируют выделение медиатора, но в этих рецепторах выделение одного медиатора регулирует другой медиатор или нейромодулятор. Большинство рецепторов – это мембраносвязанные олигомерные белки, которые связывают лиганд (нейромедиатор) с высоким сродством и высокой селективностью. В результате этого взаимодействия запускается каскад внутриклеточных изменений. Рецепторы характеризуются сродством к лиганду, количеством, насыщаемостью и способностью к диссоциации рецептор-лигандного комплекса. У некоторых рецепторов обнаружены изоформы, которые различаются сродством к определенным лигандам. Эти изоформы могут находиться в одной и той же ткани. |