фзл. Общая физиология возбудимых тканей
 Скачать 1.43 Mb. Скачать 1.43 Mb.
|

|
Лиганды - это вещества, избирательно взаимодействующие с данным рецептором. Если фармакологическое вещество активирует данный рецептор, оно является агонистом для него, а если снижает его активность – то антагонистом. Связывание лиганда с рецептором приводит к изменению конформации рецептора, вседствие чего или открываются ионные каналы, или запускается каскад реакций, приводящих к изменениям метаболизма. Передача нервных импульсов через синапсы происходит химическим путем - с помощью нейромедиаторов (нейротрансмиттеров) . В настоящее время известны следующие вещества, выполняющие медиаторные функции: ацетилхолин , катехоламины ( адреналин , норадреналин,дофамин), аминокислоты ( гамма-аминомасляная кислота , глутаминовая кислота , глицин) ,гистамин , нейроактивные пептиды . К числу самых важных нейромедиаторов мозга относятсяацетилхолин, норадреналин, серотонин, дофамин, глутамат, ГАМК, эндорфины и энкефалины. Нейротрансмиттеры являются первичными мессенджерами, но их высвобождение и определение в химических синапсах сильно отличается от эндокринных сигналов. В пресинаптической клетке, везикулы, содержащие нейротрансмиттер, высвобождают собственное содержимое локально в очень маленький объем синаптической щели. Высвобожденный трансмиттер затем диффундирует через щель и связывается с рецепторами на постсинаптической мембране. Диффузия является медленным процессом, но пересечение такой короткой дистанции, которая разделяет пре- и постсинаптические нейроны (0,1 мкм или меньше), происходит достаточно быстро и позволяет осуществлять быстрые коммуникации между нервами или между нервом и мышцей. Нейротрансмиттеры включают несколько семейств, ( ацетилхолин, ГАМК, допамин ) и (вазопрессин, брадикинин). В зависимости от их функции нейромедиаторы можно разделить на два типа: Возбуждающие - этот тип нейромедиаторов оказывает возбуждающее воздействие на нейрон. Они увеличивают вероятность того, что нейрон будет генерировать потенциал действия. К основным возбуждающим нейротрансмиттерам причисляют адреналин и норадреналин. Ингибирующие - эти нейротрансмиттеры оказывают ингибирующее действие на нейрон; они уменьшают вероятность того, что будет выработан потенциал действия. Основными нейромедиаторами ингибирующего типа считаются серотонин и гамма-аминомасляная кислота (или ГАМК). Некоторые нейротрансмиттеры, такие как ацетилхолин и дофамин, могут оказывать возбуждающий и подавляющий эффект в зависимости от типа рецепторов, которыми обладает постсинаптический нейрон. Также любой из нейромедиаторов можно отнести к одному из шести типов: 1. Ацетилхолин 2. Аминокислоты: ГАМК, глицин, глутамат, аспартат. 3. Нейропептиды: окситоцин, эндорфины, вазопрессин и др. 4. Моноамины: адреналин, норадреналин, гистамин, дофамин и серотонина. 5. Пурины: аденозин, аденозинтрифосфат (АТФ). 6. Липиды и газы: оксид азота, каннабиноиды. ГАМК – основной тормозный нейромедиатор ЦНС. У млекопитающих этот нейромедиатор встречается практически во всех отделах ЦНС, но в наибольшем количестве в синапсах головного мозга. Вторым важнейшим нейромедиатором, осуществляющим торможение в ЦНС, является глицин. В качестве нейромедиатора он наиболее распространен в филогенетически древних областях: синапсах ствола головного мозга, продолговатого и 3 спинного мозга, хотя присутствие глицина показано и в высших отделах, таких как таламус, кора мозжечка, гипоталамус, стриатум, кора головного мозга Ионотропный глициновый рецептор является первым рецептором нейромедиаторов, который был выделен из нервных клеток млекопитающих. Мутации генов субъединиц глицинового рецептора у мышей и человека приводят к возникновению нарушения поведения и двигательной функции, что подчеркивает большое значение глицинергической синаптической сигнализации. ГАМКА-рецептор, также как и глициновый, является ионотропным. Мутации в генах, кодирующих субъединицы ГАМКА-рецепторов приводят к возникновению различных типов эпилепсии, а также к развитию неврологических и психических нарушений, таких как депрессии и аффективные расстройства. Помимо ГАМКА рецепторов, в ЦНС существуют два других типа рецепторов ГАМК: метаботропные ГАМКВ и ионотропные ГАМКС. Активация глициновых и ГАМКА-рецепторов приводит к открытию каналов, проницаемых для ионов хлора, что у взрослых особей приводит к вхождению этих ионов внутрь клетки и гиперполяризации мембраны. Глициновые и ГАМКА-рецепторы могут быть локализованы как постсинаптически, так и пресинаптически. В случае пресинаптической локализации, они участвуют в регуляции высвобождения нейромедиатора. ГАМКА-рецепторы, локализованные на пресинаптических терминалях в спинном мозгу крысы, могут регулировать высвобождение глицина. Виды мышц. Механизм сокращения и расслабления скелетных мышц (теория скольжения). Сила, работа и утомление мышц. Особенности сокращения гладких мышц. По морфологическим признакам выделяют три группы мышц: 1) поперечно-полосатые мышцы (скелетные мышцы); 2) гладкие мышцы; 3) сердечную мышцу (или миокард). Механизм мышечного сокращения. В процессе сокращения мышечного волокна в нем происходят следующие преобразования: А. Электрохимическое преобразование: 1. Генерация ПД. 2. Распространение ПД по Т-системе. 3. Электрическая стимуляция зоны контакта Т-системы и саркоплазматического ретикулума, активация ферментов, образование инозитолтрифосфата, повышение внутриклеточной концентрации ионов Са2+. Б. Хемомеханическое преобразование: 4. Взаимодействие ионов Са2+ с тропонином, освобождение активных центров на актиновых филаментах. 5. Взаимодействие миозиновой головки с актином, вращение головки и развитие эластической тяги. 6. Скольжение нитей актина и миозина относительно друг друга, уменьшение размера саркомера, развитие напряжения или укорочение мышечного волокна. Передача возбуждения с двигательного мотонейрона на мышечное волокно происходит с помощью медиатора ацетилхолина (АХ). Взаимодействие АХ с холинорецептором концевой пластинки приводит к активации АХ-чувствительных каналов и появлению потенциала концевой пластинки, который может достигать 60 мВ. При этом область концевой пластинки становится источником раздражающего тока для мембраны мышечного волокна и на участках клеточной мембраны, прилегающих к концевой пластинке, возникает ПД, который распространяется в обе стороны со скоростью примерно 3—5 м/с при температуре 36 oС. Таким образом, генерация ПД является первым этапом мышечного сокращения. Вторым этапом является распространение ПД внутрь мышечного волокна по поперечной системе трубочек, которая служит связующим звеном между поверхностной мембраной и сократительным аппаратом мышечного волокна. Т-система тесно контактирует с терминальными цистернами саркоплазматической сети двух соседних саркомеров. Электрическая стимуляция места контакта приводит к активации ферментов, расположенных в месте контакта и образованию инозитолтрифосфата. Инозитолтрифосфат активирует кальциевые каналы мембран терминальных цистерн, что приводит к выходу ионов Са2+ из цистерн и повышению внутриклеточной концентрации Са2+ с 107до 105 M. Совокупность процессов, приводящих к повышению внутриклеточной концентрации Са2+ составляет сущность третьего этапа мышечного сокращения. Таким образом, на первых этапах происходит преобразование электрического сигнала ПД в химический — повышение внутриклеточной концентрации Са2+, т. е. электрохимическое преобразование. При повышении внутриклеточной концентрации ионов Са2+ тропомиозин смещается в желобок между нитями актина, при этом на актиновых нитях открываются участки, с которыми могут взаимодействовать поперечные мостики миозина. Это смещение тропомиозина обусловлено изменением конформации молекулы белка тропонина при связывании Са2+ . Следовательно, участие ионов Са2+ в механизме взаимодействия актина и миозина опосредовано через тропонин и тропомиозин. четвертым этапом электромеханического сопряжения является взаимодействие кальция с тропонином. пятым этапом электромеханического сопряжения является присоединение головки поперечного мостика к актиновому филаменту к первому из нескольких последовательно расположенных стабильных центров. При этом миозиновая головка поворачивается вокруг своей оси, поскольку имеет несколько активных центров, которые последовательно взаимодействуют с соответствующими центрами на актиновом филаменте. Вращение головки приводит к увеличению упругой эластической тяги шейки поперечного мостика и увеличению напряжения. В каждый конкретный момент в процессе развития сокращения одна часть головок поперечных мостиков находится в соединении с актиновым филаментом, другая свободна, т. е. существует последовательность их взаимодействия с актиновым филаментом. Это обеспечивает плавность процесса сокращения. На четвертом и пятом этапах происходит хемомеханическое преобразование. Последовательная реакция соединения и разъединения головок поперечных мостиков с актиновым филаментом приводит к скольжению тонких и толстых нитей относительно друг друга и уменьшению размеров саркомера и общей длины мышцы, что является шестым этапом. Совокупность описанных процессов составляет сущность теории скольжения нитей Первоначально полагали, что ионы Са2+ служат кофактором АТФазной активности миозина. Дальнейшие исследования опровергли это предположение. У покоящейся мышцы актин и миозин практически не обладают АТФазной активностью. Присоединение головки миозина к актину приводит к тому, что головка приобретает АТФазную активность. В каждом цикле соединения и разъединения головки миозина с актином расщепляется одна молекула АТФ на каждый мостик. Быстрота вращения определяется скоростью расщепления АТФ. Таким образом, в процессе хемомеханического преобразования АТФ обеспечивает разъединение головки миозина и актинового филамента и энергетику для дальнейшего взаимодействия головки миозина с другим участком актинового филамента. Эти реакции возможны при концентрации кальция выше 106М. Описанные механизмы укорочения мышечного волокна позволяют предположить, что для расслабления в первую очередь необходимо понижение концентрации ионов Са2+. Экспериментально было доказано, что саркоплазматическая сеть имеет специальный механизм — кальциевый насос, который активно возвращает кальций в цистерны. Активация кальциевого насоса осуществляется неорганическим фосфатом, который образуется при гидролизе АТФ, а энергообеспечение работы кальциевого насоса также за счет энергии, образующейся при гидролизе АТФ. Таким образом, АТФ является вторым важнейшим фактором, абсолютно необходимым для процесса расслабления. Некоторое время после смерти мышцы остаются мягкими вследствие прекращения тонического влияния мотонейронов. Затем концентрация АТФ снижается ниже критического уровня и возможность разъединения головки миозина с актиновым филаментом исчезает. Сила сокращения скелетной мышцы определяется 2 факторами: • числом Двигательных Единиц, участвующих в сокращении; • частотой сокращения мышечных волокон. Работа скелетной мышцы совершается за счет согласованного изменения тонуса (напряжения) и длины мышцы во время сокращения. Виды работы скелетной мышцы: • динамическая преодолевающая работа совершается, когда мышца, сокращаясь, перемещает тело или его части в пространстве; • статическая (удерживающая) работа выполняется, если благодаря сокращению мышцы части тела сохраняются в определенном положении; • динамическая уступающая работа совершается, если мышца функционирует, но при этом растягивается, так как совершаемого ею усилия недостаточно, чтобы переместить или удержать части тела. Во время выполнения работы мышца может сокращаться: • изотонически – мышца укорачивается при постоянном напряжении (внешней нагрузке); изотоническое сокращение воспроизводится только в эксперименте; • изометричеки – напряжение мышцы возрастает, а ее длина не изменяется; мышца сокращается изометрически при совершении статической работы; • ауксотонически – напряжение мышцы изменяется по мере ее укорочения; ауксотоническое сокращение выполняется при динамической преодолевающей работе. Утомление – физиологическое состояние мышцы, которое развивается после совершения длительной работы и проявляется снижением амплитуды сокращений, удлинением латентного периода сокращения и фазы расслабления. Причинами утомления являются: истощение запаса АТФ, накопление в мышце продуктов метаболизма. Утомляемость мышцы при ритмической работе меньше, чем утомляемость синапсов. Поэтому при совершении организмом мышечной работы утомление первоначально развивается на уровне синапсов ЦНС и нейро-мышечных синапсов. Поэтому Хаксли и Хэнсон разработали теорию скольжения нитей. Согласно ей мышца укорачивается в результате движения тонких актиновых нитей в промежутки между миозиновыми. Это приводит к укорочению каждого саркомера, образующего миофибриллы. Скольжение же нитей обусловлено тем, что при переходе в активное состояние головки отростков миозина связываются с центрами актиновых нитей и вызывают их движение относительно себя (гребковые движения). Но это последний этап всего сократительного механизма. Скрытый (латентный) период возбуждения у гладких мышц больше, чем у поперечнополосатых. У гладких мышц и пороговое раздражение выше, следовательно, возбудимость у них ниже. Сокращение гладких мышц происходит, медленнее и более продолжительно, чем поперечнополосатых мышц. Гладкие мышцы могут находиться в состоянии длительного сокращения, но оно не является тетанусом, который характерен для поперечнополосатых мышц. При длительном, или тоническом, сокращении гладких мышц в отличие от тетануса скелетных мышц обмен веществ изменяется незначительно. Следует отметить, что обмен веществ в гладких мышцах вообще менее интенсивен, чем в поперечнополосатых. Гладкие мышцы по сравнению с поперечнополосатыми обладают большей растяжимостью, что имеет существенное значение в функции органов, объем которых резко изменяется (мочевой пузырь, матка). От сокращения гладких мышц зависит не только объем полых внутренних органов, но также и перемещение их содержимого (например, пищевых масс в пищеварительном канале). Виды сокращения скелетных мышц. Механизм развития одиночных и суммированных (тетанических) сокращений мышц. Механизмы мышечного тонуса. Оптимум и пессимум мышечного сокращения. Существует два вида мышечных сокращений – одиночное и тетаническое. Одиночное мышечное сокращение является единственным видом сокращений для сердечной мышцы, а в скелетной мускулатуре оно носит искусственную этиологию и возникает в ответ на одиночный электрический сигнал и возникновение потенциала действия (ПД). Такое сокращение, длящееся » 100 мс, имеет форму волны и включает три фазы: 1 – латентный период (от 2-3 до 10 мс), длящийся от момента нанесения раздражения до начала сокращения, 2 – фаза укорочения или сокращения (40-50 мс) 3 – фаза расслабления (около 50мс). В естественных условиях импульсы поступают не одиночно, а сериями не менее 15-50 имп/с, на что мышца отвечает возникновением тетанического сокращения (тетануса). В его основе лежит явление суммации нескольких одиночных сокращений. В зависимости от частоты импульсов различают зубчатый и гладкий тетанус Зубчатый тетанус (неполный) возникает в том случае, когда каждый последующий импульс приходит в фазу расслабления мышцы. Если частота раздражения выше, и каждый последующий импульс приходит в фазу укорочения мышцы, то происходит полная суммация, и тетаническое сокращение носит слитный характер –гладкий тетанус (полный). 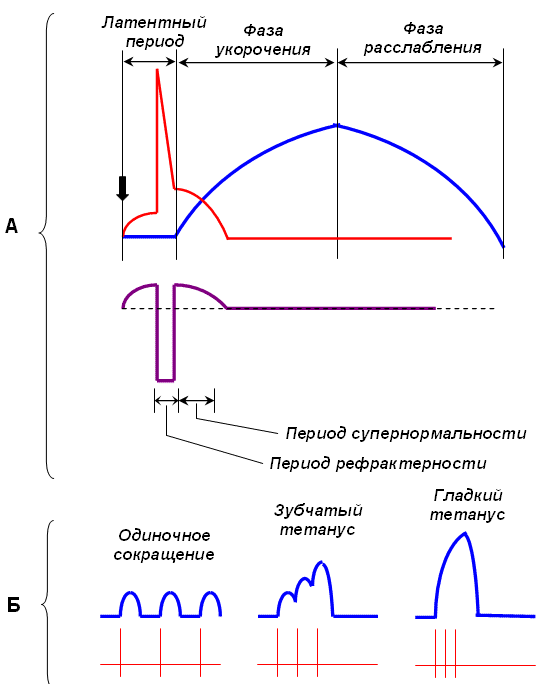 Одиночное сокращение. Раздражение мышцы или иннервирующего ее двигательного нерва одиночным стимулом вызывает одиночное сокращение мышцы. В нем различают две основные фазы: фазу сокращения и фазу расслабления. Сокращение мышечного волокна начинается уже во время восходящей ветви ПД. Длительность сокращения в каждой точке мышечного волокна в десятки раз превышает продолжительность ПД. Поэтому наступает момент, когда ПД прошел вдоль всего волокна и закончился, волна же сокращения охватила все волокно и оно продолжает быть укороченным. Это соответствует моменту максимального укорочения или напряжения мышечного волокна. Сокращение каждого отдельного мышечного волокна при одиночных сокращениях подчиняется закону "все или ничего" - сокращение, возникающее как при пороговом, так и при сверхпороговом раздражении, имеет максимальную амплитуду. Величина же одиночного сокращения всей мышцы зависит от силы раздражения. При пороговом раздражении сокращение ее едва заметно, с увеличением же силы раздражения оно нарастает, пока не достигнет известной высоты, после чего уже остается неизменной (максимальное сокращение). Это объясняется тем, что возбудимость отдельных мышечных волокон неодинакова, и поэтому только часть их возбуждается при слабом раздражении. При максимальном сокращении они возбуждены все. Суммация сокращений и тетанус. Если на мышцу действуют два быстро следующих друг за другом сильных одиночных раздражения, то возникающее сокращение будет иметь большую амплитуду, чем максимальное одиночное сокращение. Сократительные эффекты, вызванные первым и вторым раздражением, как бы складываются. Это явление носит название суммации сокращений. Для возникновения суммации необходимо, чтобы интервал между раздражениями имел определенную длительность - он должен быть длиннее рефрактерного периода, но короче всей длительности одиночного сокращения, чтобы второе раздражение подействовало на мышцу раньше, чем она успеет расслабиться. Если второе раздражение поступает, когда мышца уже начала расслабляться, на миографической кривой вершина второго сокращения будет отделяться от первого западением. Если же второе раздражение действует, когда первое сокращение еще не дошло до своей вершины, то второе сокращение как бы сливается с первым, образуя вместе с ним единую суммированную вершину. Как при полной, так и при неполной суммации ПД не суммируются. Такое суммированное сокращение в ответ на ритмические раздражения называются тетанусом. В зависимости от частоты раздражения он бывает зубчатый и гладкий. Причина суммации сокращений при тетанусе кроется в накоплении ионов Са++ в межфибриллярном пространстве до концентрации 5*106 мМ/л. После достижения этой величины дальнейшее накопление Са++ не приводит к увеличению амплитуды тетануса. После прекращения тетанического раздражения волокна вначале расслабляются не полностью, и их исходная длина восстанавливается лишь по истечении некоторого времени. Это явление называется посттетанической , или остаточной контрактурой. Она связана с тем. что требуется больше времени для удаления из межфибриллярного пространства всего Са++, попавшего туда при ритмических стимулах и не успевшего полностью удалиться в цистерны саркоплазматического ретикулюма работой Са-насосов. Если после достижения гладкого тетануса еще больше увеличивать частоту раздражения, то мышца при какой-то частоте начинает вдруг расслабляться. Это явление называется пессимумом. Он наступает тогда, когда каждый следующий импульс попадает в рефрактерность от предыдущего. Повышение частоты и силы раздражения до известного предела вызывает увеличение высоты тетанического сокращения скелетной мышцы. Наиболее благоприятная частота нервных импульсов, поступающих в скелетную мышцу, вызывает наибольшую высоту тетануса. Эта частота называется оптимальной, или оптимумом частоты. Оптимуму частоты соответствует такая частота, при которой каждое последующее раздражение застает скелетную мышцу в состоянии наибольшей возбудимости, наблюдающейся в экзальтационной фазе. |