микробиология. Общая вирусология. Механизмы противовирусной защиты. Микробиология натуральная оспа.
 Скачать 2.52 Mb. Скачать 2.52 Mb.
|

|
Модуль 7 Тема: «Общая вирусология. Механизмы противовирусной защиты. Микробиология натуральная оспа.» 1.Морфология и физиология вирусов. Вирусы – микроорганизмы, составляющие царство Vira. Отличительные признаки: 1) содержат лишь один тип нуклеиновой кислоты (РНК или ДНК); 2) не имеют собственных белоксинтезирующих и энергетических систем; 3) не имеют клеточной организации; 4) обладают дизъюнктивным (разобщенным) способом репродукции (синтез белков и нуклеиновых кислот происходит в разных местах и в разное время); 5) облигатный паразитизм вирусов реализуется на генетическом уровне; 6) вирусы проходят через бактериальные фильтры. Вирусы могут существовать в двух формах: внеклеточной (вириона) и внутриклеточной (вируса). По форме вирионы могут быть: 1) округлыми; 2) палочковидными; 3) в виде правильных многоугольников; 4) нитевидными и др. Размеры их колеблются от 15–18 до 300–400 нм. В центре вириона – вирусная нуклеиновая кислота, покрытая белковой оболочкой – капсидом, который имеет строго упорядоченную структуру. Капсидная оболочка построена из капсомеров. Нуклеиновая кислота и капсидная оболочка составляют нуклеокапсид. Нуклеокапсид сложноорганизованных вирионов покрыт внешней оболочкой – суперкапсидом, которая может включать в себя множество функционально различных липидных, белковых, углеводных структур. Строение ДНК– и РНК-вирусов принципиально не отличается от НК других микроорганизмов. У некоторых вирусов в ДНК встречается урацил. ДНК может быть: 1) двухцепочечной; 2) одноцепочечной; 3) кольцевой; 4) двухцепочечной, но с одной более короткой цепью; 5) двухцепочечной, но с одной непрерывной, а с другой фрагментированной цепями. РНК может быть: 1) однонитевой; 2) линейной двухнитевой; 3) линейной фрагментированной; 4) кольцевой; 5) содержащей две одинаковые однонитевые РНК. Вирусные белки подразделяют на: 1) геномные – нуклеопротеиды. Обеспечивают репликацию вирусных нуклеиновых кислот и процессы репродукции вируса. Это ферменты, за счет которых происходит увеличение количества копий материнской молекулы, или белки, с помощью которых на матрице нуклеиновой кислоты синтезируются молекулы, обеспечивающие реализацию генетической информации; 2) белки капсидной оболочки – простые белки, обладающие способностью к самосборке. Они складываются в геометрически правильные структуры, в которых различают несколько типов симметрии: спиральный, кубический (образуют правильные многоугольники, число граней строго постоянно) или смешанный; 3) белки суперкапсидной оболочки – это сложные белки, разнообразные по функции. За счет них происходит взаимодействие вирусов с чувствительной клеткой. Выполняют защитную и рецепторную функции. Среди белков суперкапсидной оболочки выделяют: а) якорные белки (одним концом они располагаются на поверхности, а другим уходят в глубину; обеспечивают контакт вириона с клеткой); б) ферменты (могут разрушать мембраны); в) гемагглютинины (вызывают гемагглютинацию); г) элементы клетки хозяина. 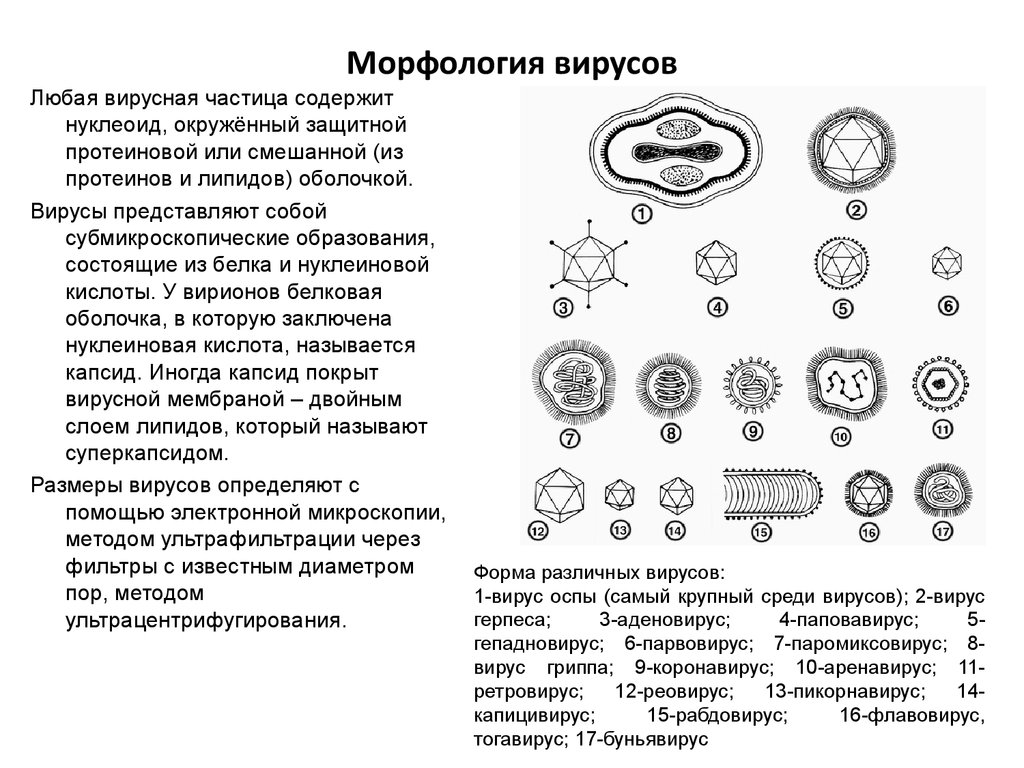 Физиология вирусов Вирус является облигатным внутриклеточным паразитом и для размножения ему требуется живая клетка. Различают три типа взаимодействия вируса с клеткой: продуктивный, или цитоцидный тип, при котором в зараженных клетках образуется новое поколение вирионов; абортивный тип, характеризующийся прерыванием инфекционного процесса в клетке, поэтому новые вирионы не образуются; интегративный тип, или вирогения, заключающийся в интеграции, т.е. встраивании вирусной ДНК в виде провируса в хромосому клетки и их совместном сосуществовании. Продуктивный тип взаимодействия вируса с клеткой осуществляется в результате размножения, т.е. репродукции вируса (от англ. reproduce – воспроизводить). Репродукция вируса происходит в несколько стадий, различающихся у разных вирусов: адсорбция вирионов на клетке; проникновение вирусов в клетку; депротеинизация или «раздевание» вирусов и высвобождение вирусного генома; биосинтез компонентов вируса; формирование вирусной частицы; выход вирионов из клетки. Вирусное инфицирование клетки начинается с адсорбции вируса на ее поверхности (рис. 55). Адсорбция вируса обеспечивается взаимодействием его поверхностных белков со специфическими рецепторами чувствительных клеток. Соответствие клеточных рецепторов и поверхностных вирусных белков определяет тропизм вируса (греч. tropos – поворот, направление), то есть способность избирательно поражать определенные клетки. Вирусы, репродуцирующиеся в клетках печени, называются гепатотропными, в клетках нервной системы – нейротропными и т.д. Проникновение вируса в клетку происходит либо путем виропексиса (рецепторного эндоцитоза), либо слияния оболочки вируса с клеточной мембраной (при наличии белка слияния), или в результате сочетания этих двух механизмов. В процессе проникновения вириона в клетку при участии клеточных ферментов происходит его депротеинизация, в результате которой удаляются поверхностные структуры вируса, и высвобождается его внутренний компонент (сердцевина, нуклеокапсид, нуклеиновая кислота). Биосинтез вирусных компонентов осуществляется в разных частях клетки, поэтому называется дизъюнктивным (от лат. disjunctus – разобщенный). Белки вируса синтезируются в результате транскрипции, т.е. «переписывания» информации с генома вируса на информационную РНК (иРНК) и последующей трансляции (считывание иРНК на рибосомах) с образованием белка вируса. Вирусная нуклеиновая кислота кодирует синтез структурных и неструктурных белков вируса. Структурные белки входят в состав вириона, а неструктурные – являются ферментами и обеспечивают репродукцию вируса. Одновременно с синтезом белка в клетке происходит и репликация (от лат. replicatio – повторение), т.е. синтез вирусных нуклеиновых кислот. Формирование вирионов происходит путем самосборки: составные части вириона транспортируются в места сборки вируса в ядре или цитоплазме. Сборка компонентов вириона происходит за счет гидрофобных, ионных, водородных связей и стерического соответствия. В результате самосборки капсомеров, образовавшихся из вирусных полипептидов, и взаимодействия их с нуклеиновыми кислотами вируса образуются нуклеокапсиды. Суперкапсидная оболочка сложноорганизованных вирусов включает в себя кроме вирусспецифических белков еще компоненты мембраны клетки. 2.Особенности патогенеза вирусных инфекций и механизмы противовирусного иммунитета. 1. Способность многих РНК и ДНК-вирусов вызывать интегративную инфекцию, которая происходит при встраивании вирусной нуклеиновой кислотой в хромосому клетки хозяина. 2. Стадия вирусемии, вирус циркулирует в крови, на лейкоцитах и проникают в кровеносные капилляры. 3. Поражение клеток иммунной системы. Угнетают Т- И В-лимфоциты. 4. Образования внутриядерных и внутриплазмотических включений, имеют диагностическое значение. Острая - инфекция сопровождается репродукцией вирусов в клетки хозяина и сравнительно быстро выделяется из организма. Очаговые и генерализованые. Персистирующая– длительное прибывание вируса в организме. Латентная, хроническая и медленной. Противовирусный иммунитет Вирусы проникают в организм через кожу или слизистые оболочки. Многие из них непосредственно поражают слизистые оболочки дыхательного и желудочно-кишечного трактов: риновирусы, коронавирусы, вирусы парагриппа, респираторно-синцитиальный вирус, ротавирусы. Другие, размножаясь в слизистой оболочке, затем быстро распространяются по крови, лимфе, нейронам: пикорнавирусы, вирусы кори, паротита, простого герпеса, гепатитов и др. Некоторые – путем переноса насекомыми и другими способами попадают в кровь и органы: альфавирусы, флавивирусы, буньявирусы и др. Противовирусный иммунитет – состояние устойчивости организма к патогенному вирусу, осуществляемое системой иммунитета. Однако кроме системы иммунитета невосприимчивость к инфекции зависит от неиммунитетных факторов. Врожденная резистентность и иммунитет Резистентность и иммунитет к вирусам зависят от комплекса причин и факторов. Существует генетически обусловленная, врожденная, неспецифическая резистентность к вирусной инфекции у одних видов по сравнению с другими видами. Животные не восприимчивы ко многим инфекциям людей и, наоборот, человек не болеет чумой собак, а последние – гриппом, ВИЧ-инфекцией, другими инфекциями человека. Такая резистентность – обычно результат отсутствия условий у данного вида для паразитирования конкретного вируса. Часто она зависит от того, что на клетках этого вида не экспрессируются рецепторы, связывающие вирус. Например, для проникновения ВИЧ-вируса в клетку нужна молекула-рецептор CD4, связывающая егоgр120, а также необходим корецепторCCR5. Вирус Эпштейн-Барр связывается сCD21 (CR2 рецептор комплемента), вирус кори – сCD46, широко представленной на лейкоцитах и других тканях, и т.д. Поэтому, вирусы тропны (обладают сродством) к клеткам и тканям, несущим к ним рецепторы: вирусы гепатита к клеткам печени – гепатоцитам, вирусы гриппа к эпителию верхних дыхательных путей, ВИЧ к Т-хелперам и т.д. На пути проникновения вирусов в клетку существуют различные неспецифические барьеры и факторы резистентности (табл. 3.1). Врожденная резистентность и иммунитет к вирусам
Кожа служит защитным барьером против большинства вирусов и они могут проникнуть в организм только при ее повреждении. То же самое относится к слизистым оболочкам, где на пути вирусов имеется слизь с вироцидными и вируссвязывающими факторами, которая удаляется вместе с ними. Ферменты слизи, протеазы, кислая среда желудочно-кишечного сока, желчь разрушают многие вирусы. Вирусы могут удаляться и выделяться всеми органами выделения: почками с мочой, печенью с желчью, секретами экскреторных желез, как в результате повреждения клеток, так и из-за повышения проницаемости эпителия. На эпителии слизистых оболочек имеются фагоциты (макрофаги и нейтрофилы), которые могут нейтрализовать вирусы, хотя сами могут служить для них мишенью, особенно когда они предварительно не активированы и находятся в покое. Дефензины эпителия и нейтрофилов разрушают многие вирусы. Нейтрализовать вирусы могут ЕК-клетки. Наиболее эффективны активированные (например, интерфероном) ЕК, которые появляются обычно через двое суток после проникновения вируса. ЕК разрушают клетки, пораженные вирусом, которые теряют антигены HLAIкласса и поэтому становятся «чужими». Комплемент, активированный вирионом по классическому или альтернативному пути, может повреждать его суперкапсид. Этот процесс более эффективен, если вирусные оболочки покрыты антителами и комплемент активируется образовавшимся комплексом антиген-антитело. Интерфероны, которые могут содержаться в секрете в значительном количестве, стимулируют резистентность клеток к вирусам. Альфа-интерферон и омега-интерферон обладают противовирусным и антипролиферативным, противоопухолевым действием. Гамма-интерферон усиливает синтез HLA-антигенов клетками, что приводит к ускорению процессов распознавания и переработки антигенов, активирует естественные киллеры, Т- и В-лимфоциты, антителогенез, адгезию лейкоцитов и моноцитов, фагоцитоз, внеклеточную и внутриклеточную вироцидность лейкоцитов, усиливает экспрессию Fc-рецепторов на моноцитах/макрофагах и поэтому связывание ими антител. Интерфероны блокируют репликацию вирусов в клетках. Они вырабатываются клетками, инфицированными вирусом, а также после стимуляции клеток лекарствами-интерфероногенами или вакцинами. Интерфероны видоспецифичны: человеческие не влияют на инфекции животных и наоборот. При стимуляции лейкоцитов вирусными и другими антигенами они выделяются в значительном количестве. Интерфероны-препараты применяют для лечения гепатитов, опухолей и других заболеваний. Интерфероны не блокируют проникновение вируса в клетку и их противовирусный эффект является опосредованным через изменение клеточного метаболизма. Сильным специфическим защитным фактором слизистых оболочек против проникновения вирусов служат секреторные IgA-антитела,которые, связываясь с ними, блокируют рецепторы вирусов и их способность адсорбироваться на клетках. Однако такие антитела имеются или после предварительной иммунизации, или после перенесенной инфекции, т.е. при наличиииммунологической памяти к антигенам данного вируса. В плазме крови или лимфе, куда вирусы попадают, преодолев барьеры кожи или слизистой оболочки, они могут нейтрализоваться IgM,IgG-антителами и комплементом, а возможно и Т-киллерами, если таковые имеются при наличии поствакцинного иммунитета или после перенесенной инфекции. Критическим моментом в развитии инфекции является связывание поверхностных структур вируса с мембраной клетки мишени, в котором участвуют или специальные белки и гликопротеиды-рецепторы или молекулы адгезии. Однако и после проникновения вируса в клетку у нее есть механизм защиты – блокировка его репликации, если она активирована интерфероном. 3.Натуральная оспа. Этиология, эпидемиология, патогенез, лабораторная диагностика, специфическая профилактика и терапия. Натуральная оспа - особо опасная высококонтагиозная острая инфекционная болезнь, которая вызывается вирусом, характеризуется воздушно-капельным (пылевым) путем передачи, двухволновой лихорадкой, резкой интоксикацией, этапным возникновением на коже и слизистых оболочках густого везикуло-пустулезного сыпи, после чего остаются рубцы. Эпидемиология натуральной оспы Источник и резервуар возбудителя - больной с последних дней инкубационного периода до полного выздоровления (максимальную опасность больные представляют с 3-8-го дня болезни). Механизм заражения натуральной оспы - аэрозольный. Передача возбудителя происходит воздушно-капельным или воздушно-пылевым путём. Факторы передачи: инфицированные вирусом воздух, пыль, нательное и постельное белье. Возможно заражение через конъюнктиву, повреждённую кожу; у беременных - трансплацентарное заражение плода. Эпидемическую опасность также представляют трупы умерших от оспы. Естественная восприимчивость людей достигает 95%. После перенесённого заболевания, как правило, развивается стойкий иммунитет, но возможно и повторное заболевание (у 0,1-1% переболевших). Натуральная оспа - высококонтагиозное заболевание. Высокий уровень заболеваемости с эпидемическим характером и циклическими подъёмами через каждые 6-8 лет регистрировался в странах Африки, Южной Америки и Азии. Чаще заражались дети в возрасте 1-5 лет. В эндемичных странах подъём заболеваемости отмечался в зимне-весенний период. Патогенез натуральной оспы При аэрозольном механизме заражения поражаются клетки слизистой оболочки носоглотки, трахеи, бронхов и альвеол. В течение 2-3 дней вирус накапливается в лёгких и проникает в регионарные лимфатические узлы, где активно реплицируется. По лимфатическим и кровеносным путям (первичная виремия) он попадает в селезёнку, печень и свободные макрофаги лимфатической системы, в которых размножается. Через 10 дней развивается вторичная виремия. инфицируются клетки кожи, почек, ЦНС, других внутренних органов и появляются первые признаки болезни. Характерный для вируса тропизм к клеткам кожи и слизистых оболочек приводит к развитию типичных оспенных элементов. В паренхиматозных органах развиваются изменения дистрофического характера. При геморрагической оспе поражаются сосуды с развитием ДВС.   |