Раздел 3. Организм как биологическая система
 Скачать 18.71 Mb. Скачать 18.71 Mb.
|

|
Наследственность — это способность организмов передавать свои признаки и свойства в ряду поколений. Изменчивость — свойство организмов приобретать новые признаки в течение жизни. Признаки — это любые морфологические, физиологические, биохимические и иные особенно сти организмов, по которым одни из них отличаются от других, например цвет глаз. Свойствами же называют любые функциональные особенности организмов, в основе которых лежит опреде ленный структурный признак или группа элементарных признаков. Признаки организмов можно разделить на качественные и количественные. Качественные признаки имеют два-три контрастных проявления, которые называют альтернативными призна ками, например голубой и карий цвет глаз, тогда как количественные (удойность коров, урожай ность пшеницы) не имеют четко выраженных различий. Материальным носителем наследственности является ДНК. У эукариот различают два типа наследственности: генотипическую и цитоплазматическую. Носители генотипической наследственности локализованы в ядре и далее речь пойдет именно о ней, а носителями цитоплазматической наследственности являются находящиеся в митохондриях и пластидах кольцевые молекулы ДНК. Цитоплазматическая наследственность передается в основном с яйцеклеткой, поэтому на зывается также материнской. В митохондриях клеток человека локализовано небольшое количество генов, однако их изме нение может оказывать существенное влияние на развитие организма, например приводить к раз витию слепоты или постепенному снижению подвижности. Пластиды играют не менее важную роль в жизни растений. Так, в некоторых участках листа могут присутствовать бесхлорофильные клетки, что приводит, с одной стороны, к снижению продуктивности растения, а с другой — та кие пестролистные организмы ценятся в декоративном озеленении. Воспроизводятся такие экзем пляры в основном бесполым способом, так как при половом размножении чаще получаются обыч ные зеленые растения. Методы генетики
Основные генетические понятия и символика Ген — это участок молекулы ДНК, или хромосомы, несущий информацию об определенном признаке или свойстве организма. Некоторые гены могут оказывать влияние на проявление сразу нескольких признаков. Такое явление называется плейотропией. Например, ген, обусловливающий развитие наследственного заболевания арахнодактилии (паучьи пальцы) вызывает искривление хрусталика, патологии мно гих внутренних органов. Каждый ген занимает в хромосоме строго определенное место — локус. Так как в соматиче ских клетках большинства эукариотических организмов хромосомы парные (гомологичные), то в каждой из парных хромосом находится по одной копии гена, отвечающего за определенный признак. Такие гены называются аллельными. Аллельные гены чаще всего существуют в двух вариантах — доминантном и рецессивном. До минантной называют аллель, которая проявляется вне зависимости от того, какой ген находится в другой хромосоме, и подавляет развитие признака, кодируемого рецессивным геном. Доми нантные аллели обозначаются обычно прописными буквами латинского алфавита (А, В, С и др.), а рецессивные — строчными (а,b, с и др.)- Рецессивные аллели могут проявляться только в том случае, если они занимают локусы в обеих парных хромосомах. Организм, у которого в обеих гомологичных хромосомах находятся одинаковые аллели, назы вается гомозиготным по данному гену, или гомозиготой (АА, аа, ААВВ, aabb и т. д.), а организм, у которого в обеих гомологичных хромосомах находятся разные варианты гена — доминантный и рецессивный — называется гетерозиготным по данному гену, или гетерозиготой (Аа, АаВbи т. д.). Ряд генов может иметь три и более структурных варианта, например группы крови по системе АВО кодируются тремя аллелями —IA, IB, i. Такое явление называется множественным аллелизмом. Однако даже в этом случае каждая хромосома из пары несет только одну аллель, то есть все три варианта гена у одного организма не могут быть представлены. Геном — совокупность генов, характерная для гаплоидного набора хромосом. Генотип — совокупность генов, характерная для диплоидного набора хромосом. Фенотип — совокупность признаков и свойств организма, которая является результатом вза имодействия генотипа и окружающей среды. Поскольку организмы отличаются между собой многими признаками, установить закономер ности их наследования можно только при анализе двух и более признаков в потомстве. Скрещи вание, при котором рассматривается наследование и проводится точный количественный учет потомства по одной паре альтернативных признаков, называется моногибридным, по двум па рам — дигибридным, по большему количеству признаков — полигибридным. По фенотипу особи далеко не всегда можно установить ее генотип, поскольку как гомози готный по доминантному гену организм (АА), так и гетерозиготный (Аа) будет иметь в феноти пе проявление доминантной аллели. Поэтому для проверки генотипа организма с перекрестным оплодотворением применяют анализирующее скрещивание — скрещивание, при котором орга низм с доминантным признаком скрещивается с гомозиготным по рецессивному гену. При этом гомозиготный по доминантному гену организм не будет давать расщепления в потомстве, тогда как в потомстве гетерозиготных особей наблюдается равное количество особей с доминантным и рецессивным признаками. Для записи схем скрещиваний чаще всего применяются следующие условные обозначения: Р (от лат. парента — родители) — родительские организмы; ♀ (алхимический знак Венеры — зеркало с ручкой) — материнская особь; ♂ (алхимический знак Марса — щит и копье) — отцовская особь; х — знак скрещивания; F1, F2, F3 и т. д. — гибриды первого, второго, третьего и последующих поколений; Fa — потомство от анализирующего скрещивания. Хромосомная теория наследственности Основоположник генетики Г. Мендель, равно как и его ближайшие последователи, не имели ни малейшего представления о материальной основе наследственных задатков, или генов. Однако уже в 1902-1903 годах немецкий биолог Т. Бовери и американский студент У. Сэттон независимо друг от друга предположили, что поведение хромосом при созревании клеток и оплодотворении позволяет объяснить расщепление наследственных факторов по Менделю, т. е., по их мнению, гены должны быть расположены в хромосомах. Данные предположения стали краеугольным кам нем хромосомной теории наследственности. В 1906 году английские генетики У. Бэтсон и Р. Пеннет обнаружили нарушение менделевского расщепления при скрещивании душистого горошка, а их соотечественник Л. Донкастер в экспериментах с бабочкой крыжовенной пяденицей открыл сцепленное с полом наследование. Результаты этих экспериментов явно противоречили менделевским, но если учесть, что к томувремени уже было известно о том, что количество известных признаков для экспериментальных объектов намного превышало количество хромосом, а это наводило на мысль, что каждая хромо сома несет более одного гена, а гены одной хромосомы наследуются совместно. В 1910 году начинаются эксперименты группы Т. Моргана на новом экспериментальном объ екте — плодовой мушке дрозофиле. Результаты этих экспериментов позволили к середине 20-х годов XX века сформулировать основные положения хромосомной теории наследственности, опре делить порядок расположения генов в хромосомах и расстояния между ними, т. е. составить пер вые карты хромосом. Основные положения хромосомной теории наследственности: 1) Гены расположены в хромосомах. Гены одной хромосомы наследуются совместно, или сцепленно, и называются группой сцепления. Число групп сцепления численно равно гаплоидно му набору хромосом.
Современные представления о гене и геноме В начале 40-х годов XX века Дж. Бидл и Э. Тейтум, анализируя результаты генетических ис следований, проведенных на грибе нейроспоре, пришли к выводу, что каждый ген контролирует синтез какого-либо фермента, и сформулировали принцип «один ген — один фермент». Однако уже в 1961 году Ф. Жакобу, Ж.-Л. Моно и А. Львову удалось расшифровать структуру гена кишечной палочки и исследовать регуляцию его активности. За это открытие им в 1965 году была присуждена Нобелевская премия по физиологии и медицине. В процессе исследования, кроме структурных генов, контролирующих развитие определенных признаков, им удалось выявить и регуляторные, основной функцией которых является проявле ние признаков, кодируемых другими генами. Структура прокариотического гена. Структурный ген прокариот имеет сложное строение, по скольку в его состав входят регуляторные участки и кодирующие последовательности. К регуляторным участкам относятся промотор, оператор и терминатор (рис. 3.8). Промотором называют участок гена, к которому прикрепляется фермент РНК-полимераза, обеспечивающий синтез иРНК в процессе транскрипции. С оператором, располагающимся между промотором и структурной по следовательностью, может связываться белок-репрессор, не позволяющий РНК-полимеразе начать считывание наследственной информации с кодирующей последовательности, и только его удале ние позволяет начать транскрипцию. Структура репрессора закодирована обычно в регуляторном гене, находящемся в другом участке хромосомы. Считывание информации заканчивается на участке гена, который называется терминатором. 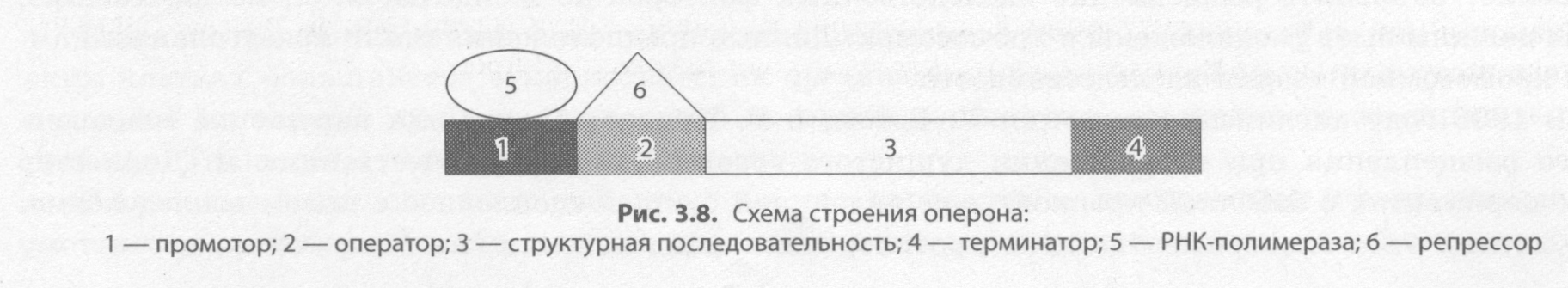 Кодирующая последовательность структурного гена содержит информацию о последователь ности аминокислот в соответствующем белке. Кодирующую последовательность у прокариот на зывают цистроном, а совокупность кодирующих и регуляторных участков гена прокариот — опероном. В целом прокариоты, к которым относится и кишечная палочка, имеют сравнительно небольшое количество генов, расположенных в единственной кольцевой хромосоме. Цитоплазма прокариот может содержать также дополнительные небольшие кольцевые или не замкнутые молекулы ДНК, которые называются плазмидами. Плазмиды способны встраиваться в хромосомы и передаваться от одной клетки к другой. Они могут нести информацию о половых признаках, патогенности и устойчивости к антибиотикам. Структура эукариотического гена. В отличие от прокариот, гены эукариот не имеют оперонной структуры, поскольку не содержат оператора, и каждый структурный ген сопровождается только промотором и терминатором. Кроме того, в генах эукариот значащие участки (экзоны) чередуются с незначащими (интронами), которые полностью переписываются на иРНК, а затем вырезаются в процессе их созревания. Биологическая роль интронов состоит в снижении вероятности мутаций в значащих участках. Регуляция генов эукариот намного сложнее, нежели описанная для про кариот. Геном человека. В каждой клетке человека в 46 хромосомах находится около 2 м ДНК, плотно упакованной в двойную спираль, которая состоит примерно из 3,2 х 109 нуклеотидных пар, что обеспечивает около 101900000000 возможных уникальных комбинаций. К концу 80-х годов XX века было известно расположение примерно 1500 генов человека, однако их общее количество оценива ли примерно в 100 тыс., поскольку только наследственных болезней у человека имеется примерно 10 тыс., не говоря уже о количестве разнообразных белков, содержащихся в клетках. В 1988 году стартовал международный проект «Геном человека», который к началу XXI века закончился полной расшифровкой последовательности нуклеотидов. Он дал возможность понять, что два разных человека на 99,9 % имеют сходные последовательности нуклеотидов, и лишь оста ющиеся 0,1% определяют нашу индивидуальность. Всего было обнаружено примерно 30-40 тыс. структурных генов, однако затем их количество было снижено до 25-30 тыс. Среди этих генов имеются не только уникальные, но и повторяющиеся сотни и тысячи раз. Тем не менее данные гены кодируют гораздо большее количество белков, например десятки тысяч защитных белков — иммуноглобулинов. 97% нашего генома является генетическим «мусором», который существует только потому, что умеет хорошо воспроизводиться (РНК, которые транскрибируются на этих участках, никогда не покидают ядро). Например, среди наших генов есть не только «человеческие» гены, но и 60% генов, похожих на гены мушки дрозофилы, а с шимпанзе нас роднит до 99 % генов. Параллельно с расшифровкой генома происходило и картирование хромосом, вследствие этого удалось не только обнаружить, но и определить расположение некоторых генов, отвечающих за развитие наследственных заболеваний, а также генов-мишеней лекарственных препаратов. Расшифровка генома человека пока не дает прямого эффекта, поскольку мы получили свое образную инструкцию по сборке такого сложного организма, как человек, но не научились из готавливать его или хотя бы исправлять погрешности в нем. Тем не менее эра молекулярной медицины уже на пороге, во всем мире идет разработка так называемых генопрепаратов, которые смогут блокировать, удалять или даже замещать патологические гены у живых людей, а не толь ко в оплодотворенной яйцеклетке. Не следует забывать и о том, что в эукариотических клетках ДНК содержится не только в ядре, но и также в митохондриях и пластидах. В отличие от ядерного генома, организация генов митохондрий и пластид имеет много общего с организацией генома прокариот. Несмотря на то что эти органеллы несут менее 1 % наследственной информации клетки и не кодируют даже полного набора белков, необходимых для их собственного функционирования, они способны существенно влиять на некоторые признаки организма. Так, пестролистность у растений хлорофитума, плюща и других наследует незначительное число потомков даже при скрещивании двух пестролистных растений. Это обусловлено тем, что пластиды и митохондрии передаются большей частью с цито плазмой яйцеклетки, поэтому такая наследственность называется материнской, или цитоплазма- тической, в отличие от генотипической, которая локализуется в ядре. 3.5. Закономерности наследственности, их цитологические основы. Моно– и дигибридное скрещивание. Закономерности наследования, установленные Г. Менделем. Сцепленное наследование признаков, нарушение сцепления генов. Законы Т. Моргана. Хромосомная теория наследственности. Генетика пола. Наследование признаков, сцепленных с полом. Генотип как целостная система. Развитие знаний о генотипе. Геном человека. Взаимодействие генов. Решение генетических задач. Составление схем скрещивания. Законы Г. Менделя и их цитологические основы. Закономерности наследственности, их цитологические основы Согласно хромосомной теории наследственности каждая пара генов локализована в паре гомо логичных хромосом, причем каждая из хромосом несет только по одному из этих факторов. Если представить, что гены являются точечными объектами на прямых — хромосомах, то схематиче ски гомозиготные особи могут быть записаны как А||А или а||а, тогда как гетерозиготная — А||а. При образовании гамет в процессе мейоза каждый из генов пары гетерозиготы окажется в одной из половых клеток (рис. 3.9). Например, если скрестить двух гетерозиготных особей, то при условии образования у каждой из них только пары гамет возможно получение всего лишь четырех дочерних организмов, три из которых будут нести хотя бы один доминантный ген А, и только один будет гомозиготен по рецес сивному гену а, т. е. закономерности наследственности носят статистический характер (рис. 3.10). В тех случаях, если гены располагаются в разных хромосомах, то при образовании гамет рас пределение между ними аллелей из данной пары гомологичных хромосом происходит совершенно независимо от распределения аллелей из других пар (рис. 3.11). Именно случайное расположение гомологичных хромосом на экваторе веретена в метафазе I мейоза и их последующее расхождение в анафазе I ведет к разнообразию рекомбинаций аллелей в гаметах. Число возможных сочетаний аллелей в мужских или женских гаметах можно определить по общей формуле 2n, где n — число хромосом, характерное для гаплоидного набора. У чело века n = 23, а возможное число со четаний составляет 223 = 8388608. Последующее объединение гамет при оплодотворении является также слу чайным, и поэтому в потомстве мож но зафиксировать независимое рас щепление по каждой паре признаков (рис. 3.11). 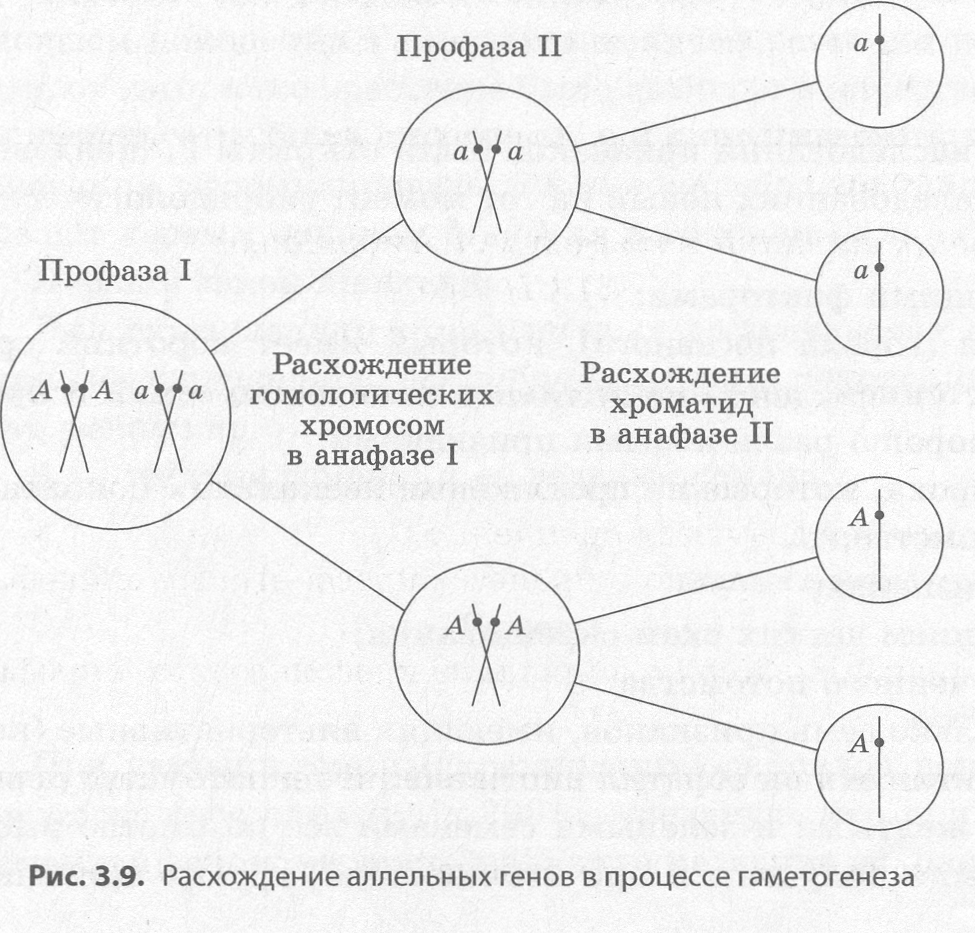 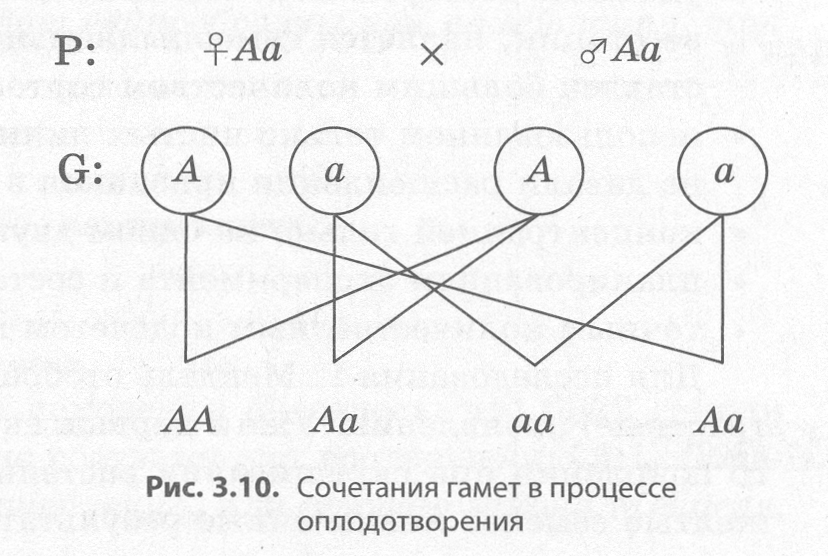 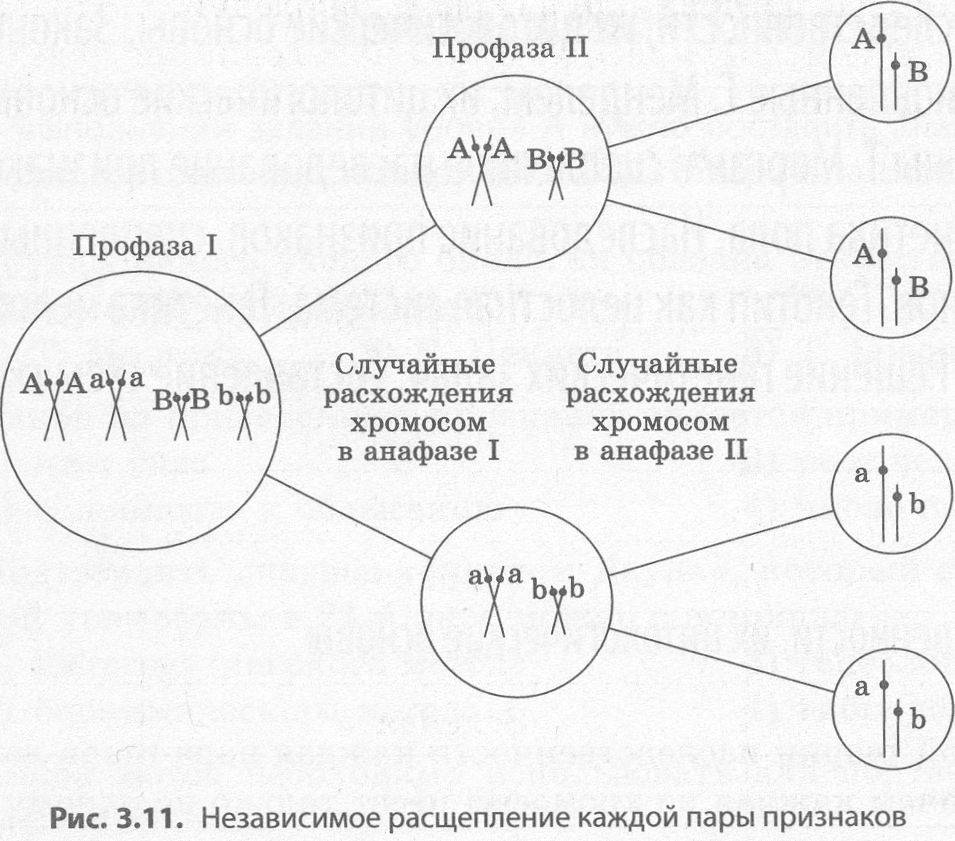 Однако число признаков у каждого организма во много раз больше числа его хромосом, ко торые можно различить под микроскопом, следовательно, каждая хромосома должна содержать множество факторов. Если представить себе, что у некоторой особи, гетерозиготной по двум парам генов, расположенных в гомологичных хромосомах, образуются гаметы, то следует учитывать не только вероятность образования гамет с исходными хромосомами, но и гамет, получивших из мененные в результате кроссинговера в профазе I мейоза хромосомы. Следовательно, в потомстве возникнут новые сочетания признаков. Данные, полученные в экспериментах на дрозофиле, лег ли в основу хромосомной теории наследственности. Другое фундаментальное подтверждение цитологической основы наследственности было полу чено при исследовании различных заболеваний. Так, у человека одна из форм рака обусловлена утратой маленького участка одной из хромосом. |