шпоры по бх-последняя итоговая. Основные системы межклеточной коммуникации эндокринная, паракринная, аутокринная регуляция
 Скачать 0.64 Mb. Скачать 0.64 Mb.
|

|
Классификация гормонов по биологическим функциям Эта классификация условна, поскольку одни и те же гормоны могут выполнять разные функции. Например, адреналин участвует в регуляции обмена жиров и углеводов и, кроме этого, регулирует частоту сердечных сокращений, АД, сокращение гладких мышц. Кортизол не только стимулирует глюконеогенез, но и вызывает задержку NaCl.
|
Гипоталамус занимает важнейшее место в иерархической системе, объединяя высшие отделы ЦНС и эндокринные железы. В клетках нейронов гипоталамуса синтезируются пептидные гормоны 2 типов. Одни через систему гипоталамо-гипофизарных сосудов поступают в переднюю долю гипофиза, где стимулируют или ингибируют синтез тропных гормонов; другие, как окситоцин и вазопрессин, поступают через аксоны нервных клеток в заднюю долю гипофиза, где они хранятся в везикулах и секрети-руются в кровь в ответ на соответствующие сигналы.
558 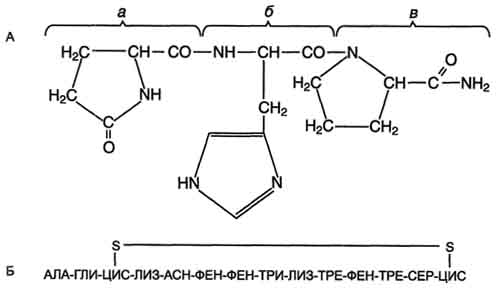 . А. Структура тиреолиберина: а - пироглутаминовая кислота; б гистидин; в - пролинамид. Б. Структура соматостатина.
Биологическое действие гормона роста. 562 Регуляция секреции гормона роста. Соматолиберин стимулирует (1), а соматостатин ингибирует (2) освобождение гормона роста (ГР) из передней доли гипофиза. ИФР-1 ингибирует секрецию соматолиберина (3) и стимулирует секрецию соматостатина (4). ИФР-1 ингибирует секрецию гормона роста также на уровне гипофиза (5). | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Гормоны задней доли гипофиза. Химическая природа. Биологическая роль. Изменения метаболизма при гипо- и гиперфункции. Задняя доля гипофиза, или нейрогипофиз, сек-ретирует 2 активных гормона - вазопрессин, или антидиуретический гормон (АДГ), и окситоцин. Окситоцин и вазопрессин - ноншепгады со сходной первичной структурой (рис. 11-16). Оба гормона образуются в гипоталамусе в нейронах разных гипоталамических ядер в форме прогормонов, из которых в результате посттрансляционной модификации образуются гормон и транспортный пептид нейрофизин (окситоцин+нейрофизин I и вазопрессин+нейрофизин II). В процессе транспорта в клетки задней доли гипофиза гормоны остаются нековалентно связанными со своими транспортными пептидами. В крови гормоны не связаны с нейрофизином. Т1/2 составляет 2-4 мин. 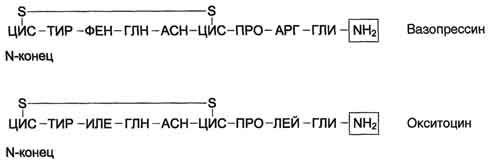 Основные биологические эффекты вазопрессина проявляются через взаимодействие с 2 типами рецепторов. V1-рецепторы расположены в клетках гладкой мускулатуры сосудов в комплексе с фосфолипазой С. Результат трансдукции сигнала в эти клетки - сокращение сосудов. V2-рецепторы расположены в клетках почечных канальцев. Взаимодействие вазопрессина с V2-рецепторами активирует аденилатциклазную систему, увеличивая в клетках концентрацию цАМФ и активность протеинкиназы . В результате этой активации происходит фосфорилирование белков, стимулирующих экспрессию генов белков, которые образуют каналы, обеспечивающие реабсорбцию воды (см. ниже подраздел VI, А). Окситоцин стимулирует сокращение гладкой мускулатуры матки, а также играет важную роль в стимуляции лактации. Он вызывает сокращение миоэпителиальных клеток молочных желёз, в результате чего происходит перераспределение молока из альвеолярных протоков в область соска. Акт сосания материнской груди стимулирует секрецию пролактина, обеспечивая образование и секрецию молока. Гипофункция может быта следствием уменьшения или полного подавления продукции тройных гормонов (пангипопитуитаризм) или частичного, при котором происходит нарушение синтеза и секреции одного или нескольких гормонов. Недостаток тропных гормонов гипофиза ведёт к резкому снижению функции периферических эндокринных желёз. Гипофизарный нанизм, или карликовость . Причина нарушения роста и физического развития - дефицит гормона роста. Большинство форм гипофизарного нанизма развивается вследствие мутаций гена гормона роста. У большинства больных гипофизарным нанизмом нарушение роста сочетается с другими эндокринными нарушениями Гиперфункция гормона роста обычно возникает в,результате образования гормонпродуцирующей опухоли соматотрофных клеток гипофиза, что приводит к повышению ростовой активности. Если гиперсекреция гормона роста возникает у детей и подростков с незакончившимся процессом окостенения эпифизар-ных хрящей, но продолжающимся ростом длинных костей, развивается гигантизм (от греч. gigantos - великан). При гигантизме увеличение костей, мягких тканей и органов происходит сравнительно пропорционально. Гиперсекреция гормона роста у взрослых людей приводит к развитию акромегалии (от греч. akros - крайний, megas - большой), при которой рост тела ускоряется, но не в длину, а в ширину с диспропорциональным увеличением размеров лица, кистей рук, стоп, черепа, увеличением размеров внутренних органов. | Гормоны щитовидной железы. Химическая природа. Биологическая роль. Изменение метаболизма при гипо- и гиперфункции. В щитовидной железе синтезируются гормоны - йодированные производные тирозина. Они объединены общим названием йодтирони-ны. К ним относят 3,5,3'-трийодтиронин (трийодтиронин, Т3) и 3,5,3',5'-тетрайодтиронин (Т4), или тироксин. Йодтиронины участвуют в регуляции многих процессов метаболизма, развития, клеточной дифференцировки, в регуляции экспрессии генов. Йодтиронины синтезируются в составе белка тиреоглобулина в фолликулах, которые представляют собой морфологическую и функциональную единицу щитовидной железы. 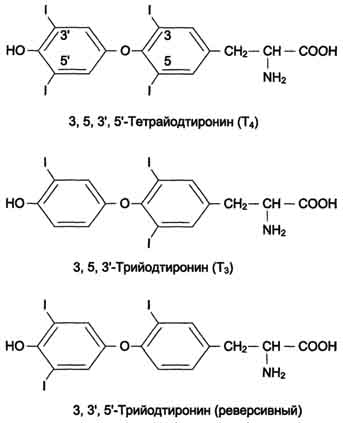 Тиреоглобулин синтезируется на рибосомах шероховатого ЭР в виде претиреоглобулина, затем переносится в цистерны ЭР, где происходит формирование вторичной и третичной структуры, включая процессы гликозилирования. Из цистерн ЭР Тиреоглобулин поступает в аппарат Гольджи, включается в состав секреторных гранул и секретируется во внеклеточный коллоид, где происходит йодирование остатков тирозина и образование йодтиронинов. Йод в виде органических и неорганических соединений поступает в ЖКТ с пищей и питьевой водой. Суточная потребность в йоде составляет 150-200 мкг. 25-30% этого количества йодидов захватывается щитовидной железой. Транспорт йодида в клетки щитовидной железы - энергозависимый процесс и происходит при участии специального транспортного белка против электрохимического градиента (соотношение концентраций I- в железе к концентрации I- в сыворотке крови в норме составляет 25:1). Работа этого йодид-переносящего белка сопряжена с Nа+,К+-АТФ-азой.Окисление I- в I+ происходит при участии гемсодержащей тиреоперокси-дазы и Н2О2 в качестве окислителя. Окисленный йод взаимодействует с остатками тирозина в молекуле тиреоглобулина. Эта реакция также катализируется тиреопероксидазой. Под действием тиреопероксидазы окисленный йод реагирует с остатками тирозина с образованием монойод-тирозинов (МИТ) и дийодтирозинов (ДИТ). Две молекулы ДИТ конденсируются с образованием йодтиронина Т4, а МИТ и ДИТ - с образованием йодтиронина Т3. Йодтиреоглобулин транспортируется из коллоида в фолликулярную клетку путём эндоцитоза и гидролизуется ферментами лизосом с освобождением Т3 и Т4. Скорость синтеза и секреции йодтиронинов регулируются гипоталамо-гипофизарной системой по механизму обратной связи. Стимулом для повышения секреции тирео-либерина и тиреотропина служит снижение концентрации йодтиронинов в крови. При физиологической концентрации йодтиронинов их действие проявляется в ускорении белкового синтеза, стимуляции процессов роста и клеточной дифференцировки. В этом отношении йодтиронины - синергисты гормона роста. Кроме того, Т3 ускоряет транскрипцию гена гормона роста. У животных при дефиците Т3 клетки гипофиза теряют способность к синтезу гормона роста. Очень высокие концентрации Т3 тормозят синтез белков и стимулируют катаболические процессы, показателем чего служит отрицательный азотистый баланс. Метаболические эффекты йодтиронинов относят в основном к энергетическому метаболизму, что проявляется в повышении поглощения клетками кислорода. Этот эффект проявляется во всех органах, кроме мозга, РЭС и гонад. В разных клетках Т3 стимулирует работу Nа+,К+-АТФ-азы, на что затрачивается значительная часть энергии, утилизируемой клеткой. В печени йодтиронины ускоряют гликолиз, синтез холестерола и синтез жёлчных кислот. В печени и жировой ткани Т3 повышает чувствительность клеток к действию адреналина и косвенно стимулирует липолиз в жировой ткани и мобилизацию гликогена в печени. Гипотиреоз у новорождённых приводит к развитию кретинизма, который проявляется множественными врождёнными нарушениями и тяжёлой необратимой задержкой умственного развития. Наиболее тяжёлые формы гипотиреоза, сопровождающиеся слизистым отёком кожи и подкожной клетчатки, обозначают термином "микседема" . Отёчность обусловлена избыточным накоплением гликозаминогликанов и воды. В подкожной клетчатке накапливается глюкуроновая и в меньшей степени хондроитинсерная кислоты. Гипотиреоз может быть также результатом недостаточного поступления йода в организм - эндемический зоб. Эндемический зоб часто встречается у людей, живущих в районах, где содержание йода в воде и почве недостаточно. Если поступление йода в организм снижается (ниже 100 мкг/сут), то уменьшается продукция йодтиронинов, что приводит к усилению секреции ТТГ (из-за ослабления действия йодтиронинов на гипофиз по механизму отрицательной обратной связи), под влиянием которого происходит компенсаторное увеличение размеров щитовидной железы (гиперплазия), но продукция йодтиронинов при этом не увеличивается. Гипертиреоз возникает вследствие повышенной продукции йодтиронинов. Диффузный токсический зоб (базедова болезнь, болезнь Грейвса) - наиболее распространённое заболевание щитовидной железы. При этом заболевании отмечают увеличение размеров щитовидной железы (зоб), повышение концентрации йодтиронинов в 2-5 раз и развитие тиреотоксикоза. Характерные признаки тиреотоксикоза: увеличение основного обмена, учащение сердцебиений, мышечная слабость, снижение массы тела (несмотря на повышенный аппетит) , потливость, повышение температуры тела, тремор и экзофтальм (пучеглазие). Гипертиреоз может возникать в результате различных причин: развитие опухоли, тиреоидит, избыточное поступление йода и йодсодер-жащих препаратов, аутоиммунные реакции. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Гормоны паращитовидных желез. Химическая природа. Биологическая роль. Изменения метаболизма при гипо- и гиперпаратиреозе. Паратгормон — вырабатывается скоплениями секреторных клеток в паренхиме железы. Необходим для поддержания концентрации ионов кальция в крови на физиологическом уровне. Снижение уровня ионизированного кальция в крови активирует секрецию паратгормона, который повышает высвобождение кальция из кости за счёт активации остеокластов. Уровень кальция в крови повышается, но кости теряют жёсткость и легко деформируются. Гормон паращитовидной железы приводит к эффектам, противоположным по действию тирокальцитонина, секретируемого С-клеткамищитовидной железы. Регуляция деятельности паращитовидных желез осуществляется по принципу обратной связи, регулирующим фактором является уровень, кальция в крови, регулирующим гормоном — паратгормон. Основная функция паратгомона заключается в поддержании постоянного уровня ионизированного кальция в крови и эту функцию он выполняет, влияя на кости, почки, и посредством витамина D — на кишечник Действие паратгормона на кости. Кость, как известно, состоит из белкового каркаса — матрикса и минераловДействие паратгормона на кость характеризуется двумя фазами. В период ранней фазы происходит увеличение метаболической активности остеокластов, это проявляется в виде выхода кальция из костей и восстановления его уровня во внеклеточной жидкости. В период поздней фазы происходит синтез белка и наблюдаются процессы образования новых клеток, а также повышается синтез лизосомальных и других ферментов, участвующих в процессах резорбции кости. Гиперкальциемия, вызванная паратгормоном, является результатом проявления обеих фаз. Механизм действия паратгормона на костную ткань осуществляется через цАМФ, активирование цАМФзависимых протеинкиназ, фосфолипазы С, диацилглицерина, инозитолтрифосфата и ионов Са. Паратгормон связывается с рецепторами, расположенными на мембранах остеокластов и остеобластов, и в клетках отмечается повышение цАМФ. Действие паратгормона на почки. Паратгормон угнетает реабсорбцию фосфатов, и в некоторой степени натрия и бикарбонатов в проксимальных канальцах почек. Это ведет к фосфатурии и гипофосфатемии. Так же увеличивается реабсорбция кальция в дистальных отделах канальцев, то есть уменьшает выделение кальция наружу. Однако при длительной гиперсекреции паратгормона развивается такая значительная гиперкальциемия, которая, несмотря на повышение реабсорбции кальция, приводит к гиперкальцийурии. Паратгормон уменьшает отложение кальция в хрусталике (при нехватке этого гормона возникает катаракта), оказывает косвенное влияние на все кальцийзависимые ферменты и катализируемые ими реакции, в том числе на реакции, формирующие свертывающую систему крови. Время биологической полужизни паратгормона составляет 8—20мин. Одним из серьёзнейших заболеваний паращитовидных желёз считается гиперактивность одного или более участков паращитовидных желёз, вызывающее выделение слишком большого количества паратгормона в кровь, что может привести к серьёзному дисбалансу кальция в организме. Такое нарушение называется гиперпаратиреоз, его последствиями могут стать гиперкальциемия и паратиреоидная остеодистрофия. | Строение, биосинтез и механизм действия кальцитриола( витамина Д3). Причины и проявления рахита. 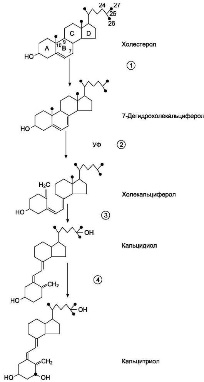 Как и другие стероидные гормоны, кальцитриол синтезируется из холестерола. Действие гормона направлено на повышение концентрации кальция в плазме крови. |