Ответы к экзамену По биохимии 1 структура и функции белков 3 ферменты 10 нуклеиновые кислоты и нуклеотиды 21
 Скачать 29.52 Mb. Скачать 29.52 Mb.
|

|
Схема иерархии регуляторных систем 1 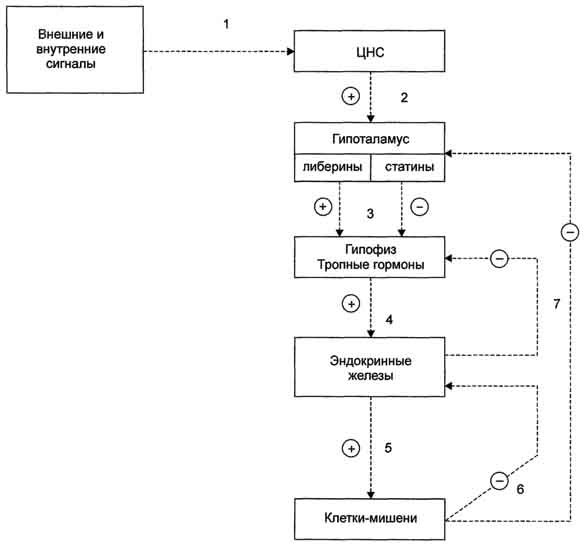 - синтез и секреция гормонов стимулируется внешними и внутренними сигналами; - синтез и секреция гормонов стимулируется внешними и внутренними сигналами; 2  ‒сигналы по нейронам поступают в гипоталамус, где стимулируют синтез и секрецию рилизинг-факторов; ‒сигналы по нейронам поступают в гипоталамус, где стимулируют синтез и секрецию рилизинг-факторов;3-рилизинг-факторы стимулируют (либерины) или ингибируют (статины) синтез и секрецию тропных гормонов гипофиза; 4‒тропные гормоны стимулируют синтез и секрецию гормонов периферических эндокринных желез; 5-гормоны эндокринных желез поступают в кровоток и взаимодействуют с клетками-мишенями; 6-изменение концентрации метаболитов в клетках-мишенях по механизму отрицательной обратной связи подавляет синтез гормонов эндокринных желез и гипоталамуса; 7-синтез и секреция тропных гормонов подавляется гормонами эндокринных желез; ⊕-стимуляция, ⊝-подавление синтеза и секреции гормонов (отрицательная обратная связь). Вопрос №135 Гормоны женских и мужских половых желез. Структура, биосинтез, регуляция, биологическое значение. Расстройства гормонообразования. Механизм действия. Мужские половые гормоны вырабатываются в основном в мужских половых железах - в интерстициальных клетках Лейдига семенников (95%). Небольшое количество андрогенов образуется в коре надпочечников. Предшественником андрогенов, как стероидных гормонов, служит холестерол, который либо поступает из плазмы в составе ЛПНП, либо синтезируется в самих железах из ацетил-КоА. Отщепление боковой цепи холестерола и образование прегненолона -скорость-лимитирующая реакция. Эта стадия стимулируется ЛГ. Тестостерон. Превращение прегненолона в тестостерон может протекать через образование прогестерона. Суточная секреция тестостерона у мужчин составляет в норме примерно 5 мг и сохраняется на протяжении всей жизни организма. Гормон циркулирует в крови в связанном с белками плазмы состоянии: альбумином (40%) и секс-гормонсвязывающим глобулином. Лишь 2% от общего количества гормона в крови транспортируется в свободном виде, и именно такие молекулы проявляют биологическую активность. Дигидротестостерон. В семенных канальцах, предстательной железе, коже, наружных половых органах тестостерон служит предшественником более активного андрогена - дигидротестостерона. Большое количество гормона - следствие периферических превращений, и суммарная суточная секреция дигидротестостерона составляет 400 мкг, что почти в 10 раз меньше уровня секреции тестостерона. В препубертатный период секреция андрогенов подавляет по механизму отрицательной обратной связи секрецию гонадотропина до начала пубертатного периода, когда гипофизарные клетки становятся менее чувствительными к ингибирующему действию циркулирующих в крови андрогенов. Эта потеря чувствительности приводит к циклически импульсному освобождению ЛГ и ФСГ. ЛГ стимулирует образование тестостерона интерстициальными клетками Лейдига, а ФСГ, связываясь с рецепторами клеток Сертоли в семенниках, стимулирует сперматогенез. К мишеням тестостерона относят сперматогонии, мышцы, кости, почки, мозг. Подобно другим стероидным гормонам, андрогены образуют внутри клетки комплекс с рецептором, который связывается с определённым участком хроматина, активируя специфические гены, белковые продукты которых опосредуют биологические эффекты андрогенов. Физиологическое действие андрогенов различно в разные периоды жизни организма. У эмбриона под действием андрогенов образуются придаток яичка, семявыносящий проток и семенной пузырёк. У плода мужского пола происходит маскулинизация мозга. Поскольку андрогены в организме обладают мощным анаболическим действием и стимулируют клеточное деление, повышенный уровень андрогенов в препубертатный период приводит к скачкообразному увеличению линейных размеров тела, увеличению скелетных мышц, росту костей, но одновременно способствуют и остановке роста, так как стимулируют сращение эпифизов длинных костей с их стволами. Андрогены вызывают изменение структуры кожи и волос, снижение тембра голоса вследствие утолщения голосовых связок и увеличения объёма гортани, стимулируют секрецию сальных желёз. Женские половые гормоны - эстрогены и прогестины, среди которых наиболее активны 17β-эстрадиол и прогестерон. В клетках теки синтезируется очень небольшое количество эстрогенов. Значительная часть эстрогенов продуцируется путём периферической ароматизации андрогенов в жёлтом теле, фетоплацентарном комплексе (во время беременности), корой надпочечников, в жировых клетках, печени, коже и других тканях, где обнаружена повышенная ароматазная активность. В клетках гранулёзы может синтезироваться менее активный эстроген - эстрон, а ещё менее активный эстриол образуется из эстрона в крови. Биологической активностью обладает только свободная форма. В детском возрасте незрелые яичники вырабатывают небольшое количество гормонов, поэтому концентрация эстрогенов в крови низкая. В пубертатный период чувствительность гипоталамо-гипофизарной системы к действию ЛГ и ФСГ снижается. Импульсная секреция гонадотропин-рилизинг-гормона устанавливает суточный ритм секреции ЛГ и ФСГ. В начале каждого менструального цикла секреция ФСГ и ЛГ вызывает развитие первичных фолликулов. Созревающий фолликул в результате совместного действия ЛГ, стимулирующего продукцию андрогенов клетками теки, и ФСГ, стимулирующего ароматизацию андрогенов, секретирует эстрогены, которые по механизму отрицательной обратной связи угнетают секрецию ФСГ. Концентрация ФСГ в крови остаётся низкой ещё и в результате торможения секреции этого гормона белком ингибином, выделяемым яичниками. По мере созревания фолликула (фолликулярная фаза) концентрация эстрадиола повышается, чувствительность гипофизарных клеток к гонадолиберину возрастает, и эстрадиол по механизму положительной обратной связи повышает секрецию ЛГ и ФСГ. Повышение секреции ЛГ приводит к овуляции - освобождению яйцеклетки из лопнувшего фолликула. После овуляции клетки гранулёзы превращаются в жёлтое тело, которое, помимо эстрадиола, начинает вырабатывать всё большее количество основного гормона лютеиновой фазы - прогестерона (прогестина). Если возникает беременность, жёлтое тело продолжает функционировать и секретировать прогестерон, однако на более поздних этапах беременности прогестерон в основном продуцируется плацентой. Если оплодотворение не происходит, высокая концентрация прогестерона в плазме крови по механизму отрицательной обратной связи угнетает активность гипоталамо-гипофизарной системы, тормозится секреция ЛГ и ФСГ, жёлтое тело разрушается, и снижается продукция стероидов яичниками. Наступает менструация, которая длится примерно 5 дней, после чего начинает формироваться новый поверхностный слой эндометрия, и возникает новый цикл. Эстрогены стимулируют развитие тканей, участвующих в размножении, определяют развитие многих женских вторичных половых признаков, регулируют транскрипцию гена рецептора прогестина. Эстрогены стимулируют синтез транспортных белков тиреоидных и половых гормонов. Эстрогены могут индуцировать синтез факторов свёртывания крови II, VII, IX и X, уменьшать концентрацию антитромбина III. Эстрогены оказывают влияние на обмен липидов. Так, увеличение скорости синтеза ЛПВП и торможение образования ЛПНП, вызываемое эстрогенами, приводит к снижению содержания холестерола в крови. Прогестерон, образующийся жёлтым телом во время менструации в лютеиновую фазу, секретируется также фетоплацентарным комплексом во время беременности. Действие прогестерона в основном направлено на репродуктивную функцию организма. Образование прогестерона отвечает за увеличение базальной температуры тела на 0,2-0,5 °С, которое происходит сразу после овуляции и сохраняется на протяжении лютеиновой фазы менструального цикла. При высоких концентрациях прогестерон взаимодействует с рецепторами, локализованными в клетках почечных канальцев, конкурируя таким образом с альдостероном. В результате конкурентного игибирования альдостерон теряет возможность стимулировать реабсорбцию натрия. Прогестерон может также оказывать действие и на ЦНС, в частности вызывать некоторые особенности поведения в предменструальный период. Вопрос №136 Репродуктивные циклы человека и их гормональная регуляция. 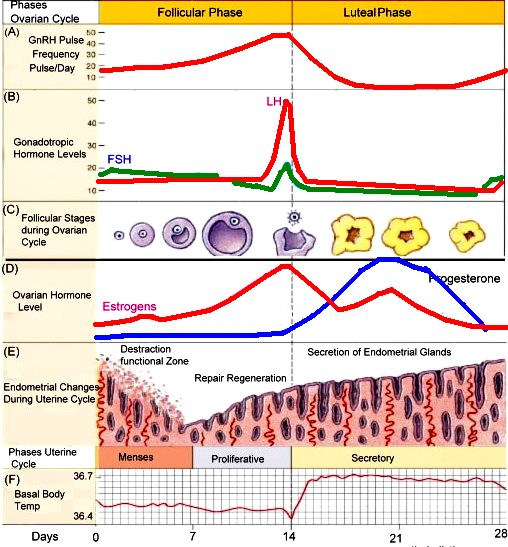 Вопрос №137 Гормоны, регулирующие обмен углеводов, жиров и аминокислот; инсулин, глюкагон, кортизол, адреналин. И 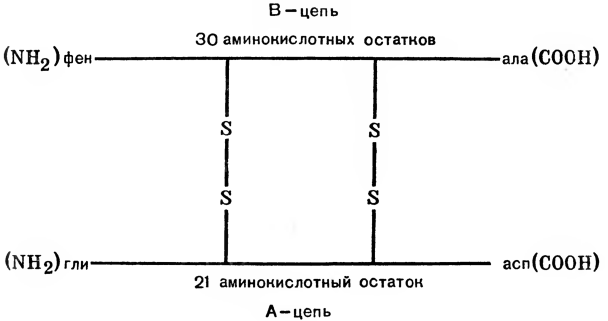 нсулин нсулинБелково-пептидной природы. Состоит из 2 ппц, соединенных дисульфидными связями. Синтезируется в β-клетках островков Лангерганса (поджелудочная железа). Синтезируется в виде неактивного предшественника. Активируется частичным протеолизом. Действует через специфические инсулиновые рецепторы: может менять активность фермента путем фосфорилирования или дефосфорилирования и/или индуцировать транскрипцию и синтез новых белков-ферментов. Влияние на обмен веществ: Углеводный:
Липидный:
Белковый:
Стимулирует синтез глюкозы. При недостатке инсулина возникает сахарный диабет.С возрастом концентрация Са2+ снижается и нарушается секреция. В крови время полужизни - 3-5 минут. После действия разрушается в печени под действием инсулиназы (расщепляет цепи инсулина). Глюкагон Состоит из 39 аминокислотных остатков. Синтезируется в α-клетках островков Лангерганса (поджелудочная железа). Действует через цАМФ, рецепторы на поверхности мембраны. Гипергликемический фактор (повышает уровень глюкозы в крови). Влияние на обмен веществ: Углеводный:
Липидный :
Адреналин Производное тирозина. Катехоламин. Синтезируется в мозговом слое надпочечников, синтез и секреция под влиянием ЦНС. Действует через цАМФ, рецепторы находятся на поверхности мембраны (α- и β-адренергические). Гормон стресса. Повышает концентрацию глюкозы в крови, т.к. активирует гликогенфосфорилазу в печени. В экстренных ситуациях активирует мобилизацию гликогена в мышечной ткани с образованием глюкозы для мышц. Тормозит секрецию инсулина. Кортизол Синтезируется из холестерола путем гидроксилирования через прегненолон и прогестерон. Синтезируется в корковом веществе надпочечников. Рецепторы в цитоплазме.
Вызывает инволюцию лимфоидной ткани, гибель лимфоцитов. Производные кортизола обладают противовоспалительной функцией (ингибируют фосфолипазу А2, что ведет к снижению уровня простагландинов - медиаторов воспаления). Гиперкортицизм. Причины:
В результате активирования глюконеогенеза, распада гликогена повышается концентрация глюкозы в крови. Возникает стероидный диабет (худые конечности, большой живот, лунообразное лицо). Вопрос №138 Изменения гормонального статуса и метаболизма при сахарном диабете. Инсулинзависимый и инсулиннезависимый сахарный диабет. Сахарный диабет - заболевание, связанное с частичным или полным отсутствием инсулина.
Биохимические проявления сахарного диабета
Осложнения сахарного диабета:
А: Острые осложнения проявляются в виде комы (нарушение обмена, потеря сознания). Виды комы, в основе которых ацидоз и дегидратация тканей: I - кето-ацидотическая кома - повышенный синтез кетоновых тел и ацидоз; II - лакто-ацидотическая кома - нарушение кровообращения, снижение функции гемоглобина, которое вызывает гипоксию. Следовательно, катаболизм глюкозы смещается в сторону "анаэробного" гликолиза до лактата. Образуется много молочной кислоты, возникает ацидоз; III - гиперосмолярная кома - из-за гипергликемии повышено осмотическое давление крови, и вода переносится из клеток в сосудистое русло, возникает дегидратация. В результате нарушается водно-электролитный обмен. Следовательно, происходит снижение периферического кровотока (мозга и почек) и гипоксия. Б: Поздние осложнения: основная причина - гипергликемия. В результате происходит неферментативное (спонтанное) гликозилирование белков, при этом нарушается их функция. Так возникают различные "-патии" (ангио-, нейро-, нейро-, ретино-). Например, в результате гликозилирования гемоглобина образуется гликозилированный ("гликированный") гемоглобин - HbA1c. В норме концентрация HbA1c - 5%. При сахарном диабете - до 50%. У него снижается сродство к кислороду → гипоксия. В хрусталике глюкоза присоединяется к кристаллину, что повышает агрегацию молекул. Следовательно, возникает помутнение хрусталика, приводящее к катаракте. При сахарном диабете нарушается синтез коллагена: из-за гликозилирования нарушается функция базальных мембран (например, кровеносных сосудов), следовательно, нарушается проницаемость сосудов и кровотока (в нижних конечностях). Это приводит к возникновению синдрома диабетической стопы и гангрене. Присоединение глюкозы к апо-белкам В100 ЛНП изменяет их строение, они захватываются макрофагами как чужеродные, проникают в поврежденный эндотелий сосудов, повышая риск атеросклероза. Лечение сахарного диабета:
Вопрос №139 Гормоны, регулирующие обмен кальция и фосфора: паратгормон, кальцитонин, калитриол. Структура, регуляция. Нарушения - гипо- и гиперпаратиреоз. Нормальная концентрация Са2+ в крови - 2,12-2,62 мМ/л. В организме содержится 100-1200 г Са2+: 99% - в минерализованных тканях (костях, зубах) в виде кристаллов гидроксиапатита. Остальная часть (1%) - в крови:
Функции Са2+ :
Гормоны: 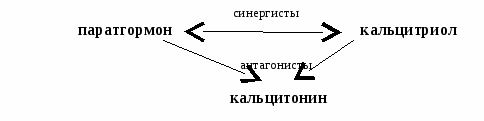 Паратгормон Состоит из 84 аминокислотных остатков. Синтезируется в клетках паращитовидной железы. Действует через цАМФ. Повышает концентрацию Са2+ в крови. Органы мишени:
Клетки-мишени - остеобласты. Они синтезируют биологически активные вещества - цитокины, которые воздействуют на остеобласты и активируют их, а остеокласты резорбируют кость. Следовательно, Са2+ выходит в кровь;
Кальцитриол Производное холестерола. Рецепторы в цитоплазме. Вторых посредников нет. В коже под УФ холестерол превращается в витамин D3 - холекальциферол. Он поступает в печень, где гидроксилируется (присоединение ОН-группы в положении 25). Затем поступает в почки, где гидроксилируется по 1 положению. Образуется 1,25-дигидроксихолекальциферол = кальцитриол. Органы-мишени:
Кальцитонин Состоит из 32 аминокислотных остатков. Вырабатывается в парафолликулярных клетках щитовидной железы. Действует через цАМФ. Понижает концентрацию Са2+ в крови. Органы-мишени:
Вопрос №140 Производные арахидоновой кислоты - эйкозаноиды. Представители, их синтез и биологическая роль. Эйкозаноиды - это большая группа веществ, которые могут синтезироваться почти всеми типами клеток, за исключением эритроцитов, и как гормоны местного действия оказывают эффекты по паракринному или аутокринному механизму через специфические рецепторы. Главные биологические эффекты эйкозаноидов: • участвуют в регуляции сокращений гладкой мускулатуры (разные типы эйкозаноидов вызывают вазоконстрикцию или вазодилатацию, бронхоконстрикцию или бронходилатацию); • регулируют экскрецию воды и Na+ почками и артериальное давление; • участвуют в развитии воспаления; • регулируют свертываемость крови Основные классы эйкозаноидов представлены: • простагландинами PG (включая простациклины); • тромбоксанами; • лейкотриенами. Исходными субстратами для синтеза эйкозаноидов являются полиеновые жирные кислоты с 20 атомами углерода. Главный субстрат для синтеза эйкозаноидов у человека - арахидоновая кислота (20:4 ω-6). Полиеновые кислоты с 20 атомами углерода поступают в организм человека с пищей или образуются из незаменимых жирных кислот с 18 атомами углерода, также поступающих с пищей. В разных тканях из арахидоновой кислоты под действием специфического для этой ткани набора ферментов образуются различные эйкозаноиды. Обычно в каждом типе клеток синтезируется преимущественно один тип эйкозаноидов. Образовавшиеся в клетке эйкозаноиды выходят из нее и взаимодействуют с рецепторами на поверхности этой же клетки (аутокринный механизм) или с рецепторами на соседних клетках (паракринный механизм). Время полураспада простагландинов равно нескольким минутам. Для каждого эйкозаноида есть несколько типов рецепторов, которые располагаются в мембране клеток рядом с аденилатциклазой, некоторые из простагландинов взаимодействуют с G-белками аденилатциклазной системы. Такие простагландины, взаимодействуя со своими рецепторами, могут модулировать активность аденилатциклазы. Ответ клетки на действие эйкозаноидов определяется типом ее рецепторов. С 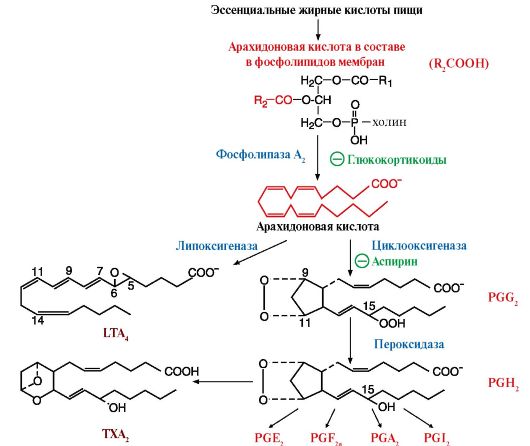 интез эйкозаноидов начинается после отделения жирной кислоты от фосфолипидов мембран под действием фермента фосфолипазы A2. Арахидоновая кислота переходит в цитозоль клетки и становится доступной для синтеза эйкозаноидов. Синтез основной группы эйкозаноидов - простагландинов, простациклинов и тромбоксанов - начинается с действия на полиеновую кислоту фермента - простагландинсинтазы. интез эйкозаноидов начинается после отделения жирной кислоты от фосфолипидов мембран под действием фермента фосфолипазы A2. Арахидоновая кислота переходит в цитозоль клетки и становится доступной для синтеза эйкозаноидов. Синтез основной группы эйкозаноидов - простагландинов, простациклинов и тромбоксанов - начинается с действия на полиеновую кислоту фермента - простагландинсинтазы. Если арахидоновая кислота подвергается действию другого фермента - липоксигеназы, то образуются молекулы с тремя сопряженными двойными связями (отсюда название «лейкотриены»). Они имеют несколько вариантов структур и в основном участвуют в развитии аллергических реакций. Синтез большинства эйкозаноидов увеличивается при воспалительных процессах. Активность фосфолипазы А2 при этих состояниях повышается и субстраты становятся доступными для синтеза эйкозаноидов. Ингибиторами синтеза эйкозаноидов являются: - глюкокортикоиды, которые индуцируют синтез группы белков, ингибирующих активность фосфолипазы А2, и таким образом подавляют синтез всех типов эйкозаноидов, участвующих в воспалении. Эти препараты обладают сильным противовоспалительным свойством. - аспирин и другие нестероидные противовоспалительные препараты необратимо ингибируют циклооксигеназу. Роль эйкозаноидов в регуляции свертывания крови. В норме свертывающая и противосвертывающая системы крови пребывают в состоянии равновесия, при котором кровь находится в жидком состоянии, но способна быстро образовывать тромб при возникновении соответствующих условий. При патологии или при действии фармакологических средств это равновесие может смещаться в любую сторону. В норме клетки эндотелия сосудов продуцируют простациклины PGI2, PGE2, PGD2, которые препятствуют агрегации тромбоцитов и сужению сосуда; TXA2, стимулирующий агрегацию тромбоцитов, в этих условиях не секретируется. ТХА2 секретируется тромбоцитами только в результате их активации - например, при контакте с поврежденной стенкой кровеносного сосуда. При разрушении клеток эндотелия (например, в результате образования атеросклеротической бляшки) синтез PGI, PGE, PGD снижается. Тромбоциты активируются в месте контакта с поврежденной стенкой сосуда и секретируют ТХА2, что стимулирует образование тромба в области повреждения эндотелия сосудов и развитие инфаркта. Если с пищей поступает больше жирных кислот ω-3, которые в большом количестве содержатся в рыбьем жире, то эти кислоты включаются преимущественно в фосфолипиды мембран (вместо арахидоновой) и после действия фосфолипазы А2 являются основными субстратами для синтеза В норме клетки эндотелия продуцируют PGI2, который вызывает релаксацию гладкой мускулатуры сосудов и ингибирует агрегацию тромбоцитов. Тромбоциты в неактивном состоянии не продуцируют тромбоксаны, кровь находится в жидком состоянии. NO - оксид азота, продуцируемый ферментом NO-синтазой, также обладает вазодилятаторным эффектом эйкозаноидов. При поражении клеток эндотелия сосуда (например, в результате развития атеросклеротической бляшки) синтез PGI2 в данном участке стенки сосуда не происходит. В это время тромбоциты контактируют с поврежденной сосудистой стенкой, в них активируется фосфолипаза А2, освобождается арахидоновая кислота и из нее синтезируется ТХА2. ТХА2 стимулирует агрегацию тромбоцитов и сокращение стенок сосуда, в результате чего на поврежденном участке сосуда образуется тромб, происходит резкое сужение просвета сосуда, нарушается кровоснабжение ткани и может развиться инфаркт Вопрос №141 Механизмы межклеточной сигнализации с помощью химических посредников и регуляторов. Внутриклеточные и внеклеточные рецепторы сигнальных молекул. Чаще всего сигналом служат растворенные во внеклеточной жидкости химические вещества. Межклеточную сигнализацию с помощью таких веществ часто подразделяют на три основных типов - аутокринную, паракринную и эндокринную. Аутокринная сигнализация - клетки отвечают на вещества, вырабатываемые этими же клетками. Паракринная сигнализация - сигнализирующие клетки влияют на рядом расположенные клетки. Дистантная сигнализация(эндокринная сигнализация)- при этом гормон или действуют на клетки-мишени вдалеке от места его выработки; у человека гормон (сигнальное вещество) в таких случаях переносится кровью. Клеточные рецепторы можно разделить на два основных класса - мембранные рецепторы и внутриклеточные рецепторы. Основные типы мембранных рецепторов:
Основные типы внутриклеточных рецепторов: Внутриклеточными рецепторами чаще всего служат белки, которые, связываясь с лигандом, выполняют роль транскрипционных факторов. Вопрос №142 Понятие о первых и вторых посредниках в межклеточной сигнализации. Циклический аденозинмонофосфат как второй посредник. Каскад цАМФ и цГМФ. Характеристика первичных посредников. 1)Физические факторы (давление, излучение). 2)Гуморальные факторы (БАВ, химические вещества) - неорганические ионы; обладающие высокой специфичностью, витамины обладающие физической активностью, гормоны 3)Кодовые сигналы. Характеристика вторичных посредников. Большая часть первичных посредников не проникает в клетку, а действует на хеморецепторы мембраны, активирует или тормозит активность аденилатциклазной системы (АЦС), гуанилат- циклазной (ГЦС), фосфолипазы С, работу Са-мембранного механизма. В итоге изменяется пассивный, активный транспорт ионов, степень связывания Са++ белков кальмодулином и др., т, е. процесс внутриклеточной передачи информации довольно универсален. Аденилатциклазный механизм Гормоны, у которых рецепторы находятся на поверхности клетки, в нее не проникают, а действуют через вторых посредников. Второй посредник - цАМФ, образуется в цитоплазме.
 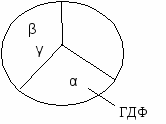
с  остоит из трех субъединиц (α, β, γ), α в неактивном виде соединена с ГДФ остоит из трех субъединиц (α, β, γ), α в неактивном виде соединена с ГДФ
М 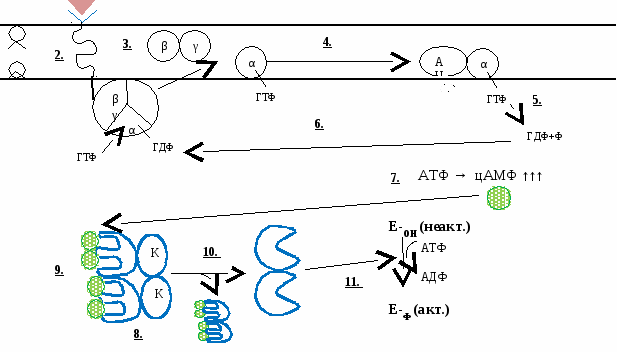 еханизм действия: еханизм действия:
Таким образом, механизм действия через вторых посредников можно представить как «каскад» - цепь последовательных биохимических реакций, приводящих к усилению сигнала. Механизм действия гормонов через рецепторы, обладающие ферментативной активностью Гормоны, у которых рецепторы находятся на поверхности клетки, в нее не проникают, а действуют через вторых посредников. Второй посредник - цГМФ. Рецептор состоит из двух частей:

Вопрос №143. Ионы кальция как вторые посредники. Каскад ионов Са2+. Са2+ как второй посредник Гормоны, у которых рецепторы находятся на поверхности клетки, в нее не проникают, а действуют через вторых посредников. И 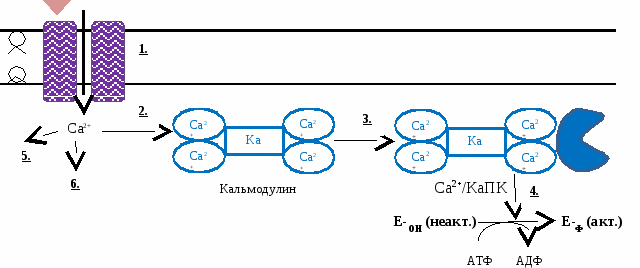 спользование Са2+ как второго посредника возможно благодаря тому, что в цитоплазме может повышаться его концентрация. спользование Са2+ как второго посредника возможно благодаря тому, что в цитоплазме может повышаться его концентрация.
Вопрос №144 Инозитол-3-фосфат как вторичный посредник. Механизм действия через липиды мембран Гормоны, у которых рецепторы находятся на поверхности клетки, в нее не проникают, а действуют через вторых посредников. Участвуют два посредника - ИФ3 (инозитолтрифосфат) и ДАГ (дтацилглицерол), которые образуются из фосфолипидов мембран (ФИФ2 - фосфатидилинозитол-4,5-бифосфат). 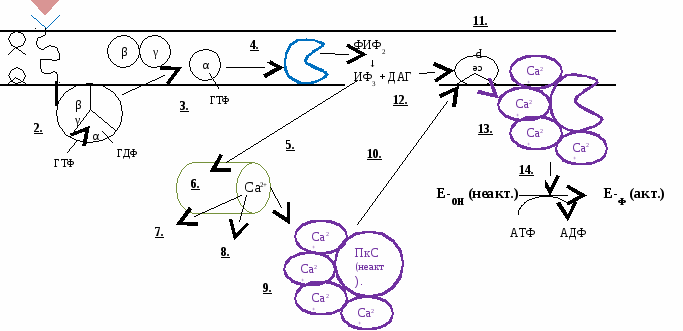

|