Савельев. Происхождение мозга
 Скачать 1.82 Mb. Скачать 1.82 Mb.
|

Глава II. Возникновение нервных клеток и мозга Причиной возникновения нервной системы стала низкая скорость получения информации о внешнем и внутреннем мире организма с донервной организацией. Его ткани состояли из клеток со сходной химической, электромагнитной и механической чувствительностью. Такой организм не мог оперативно получить дифференцированные сведения из внешнего мира и от собственного тела (Lipps, Signor, 1992). Решение проблемы без участия нервной системы было найдено растениями и одноклеточными животными, что разбиралось в главе I. Этот путь оказался биологически очень успешным и привёл к появлению процветающих групп организмов. Можно сказать, что растения и одноклеточные животные выиграли ничуть не меньше, чем животные с развитой нервной системой. Тем не менее нервная система возникла. Она давала одно, но огромное преимущество, которого невозможно достичь при помощи универсальных свойств неспециализированных клеток. Это увеличение скорости реакции организма на изменяющиеся внутренние и внешние условия. Как только нервная система позволила животным быстро адаптироваться к изменяющимся условиям, активно разыскивать пищу и половых партнёров, наступило время динамичной эволюции животного мира. Скорость решала почти все проблемы. Конкуренция за источники питания привела к появлению бесконечного разнообразия растительноядных организмов, но они сами были питательнее растений. Ещё более быстрые и эффективные хищники должны были обладать более совершенной нервной системой, чем травоядные. Это касалось как позвоночных, так и беспозвоночных животных. Кто обладал эффективной нервной системой, тот быстрее поднимался на вершину пищевой цепочки, питаясь не грубой растительной пищей, а калорийными родственниками. В самом общем виде эта ситуация сохранилась и в социальных структурах высших приматов. Преимущества и достоинства нервной системы вполне понятны. Причины её появления вполне логичны и не вызывают сомнения. Совершенно иная ситуация с вопросом о том, как это произошло. Нет ясного представления о механизмах появления первых нервных клеток. Почему они дифференцировались от других тканей и от чего возникли их уникальные свойства? Почему мозг позвоночных имеет 5 гомологичных отделов? Почему так различно поведение животных из одной систематической группы и так сходно у отдалённых видов? Как возникли и строились ассоциативные центры на различных структурных принципах? Эти 122 Происхождение нервных клеток Происхождение нервных клеток в процессе эволюции животных довольно спорно. Существует несколько основных точек зрения на эту проблему. Наибольшее распространение получила точка зрения братьев Гертвигов (Hertwig, 1878). Их гипотеза состоит в том, что нервные и мышечные клетки происходят из эктодермального эпителия. Они возникают независимо друг от друга и вступают во взаимодействия уже вторично (рис. II-1, а-в).В соответствии с гипотезой Гертвигов источником всех нервных клеток является первичная чувствительная клетка, возникшая из эктодермы и получившая возможность воспринимать раздражения, генерировать и проводить возбуждение. Специализированные отростки этих клеток вступают в связь с независимо возникшими мышечными клетками. По мнению авторов этой гипотезы, мышечные волокна возникли из клеток, сходных с эпителиально-мышечными клетками кишечнополостных. Такой простейший нервно-мышечный комплекс развивается по пути дифференциации нервного аппарата, что приводит к возникновению вставочных нейронов или ганглиев (см. рис. II-1, в). Нервная система подразделяется на чувствительные клетки, лежащие в эктодерме, и вставочные нейроны, обладающие интегративной функцией. Вставочные нейроны являются предвестниками централизации обработки сенсорной информации и возникновения мозга. На выходе расположены эффекторные нейроны, иннервирующие мышцы и железы. Другая точка зрения на происхождение нервных клеток сформировалась в работах Клейненберга и Заварзина (Kleinenberg, 1872; Заварзин, 1950). Её суть сводится к тому, что нервная и мышечная системы рассматриваются как «единое и нераздельное целое». 123 Рис. II-1. Две гипотезы происхождения нервных и мышечных клеток. а-в — гипотеза братьев Гертвигов; г-е — гипотеза Клейненберга-Заварзина. По гипотезе Гертвигов нервные и мышечные клетки происходят независимо из эктодермы, а затем объединяются в нервно-мышечный комплекс. По представлениям Клейненберга-Заварзина, из эктодермы возникает клетка одновременно с сенсорными и моторными функциями. Затем она дифференцируется на нервную и мышечную системы. Красные — мышечные клетки или волокна, синие и фиолетовые — чувствительные нервные клетки. 124 125 Рис. II-2. Актиния с симбионтом рода Amphiprion. а — микроскопический срез и его увеличенные фрагменты (б-д). На тёмном фоне в поляризованном свете видны стрекающие спироцисты (г). 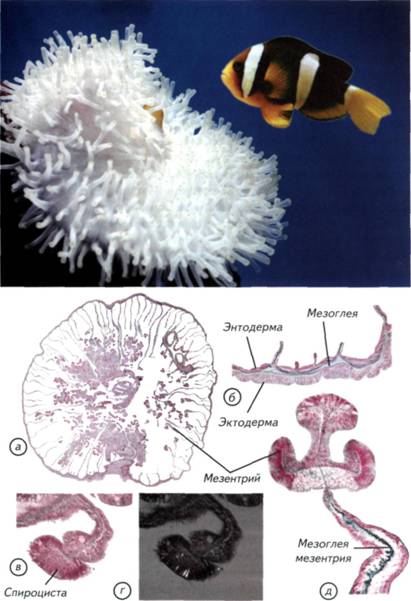 128 129 Рис. II-3. Гипотетические этапы происхождения нервных клеток в процессе эволюции многоклеточных.  а — разрез современной губки с простейшей асконоидной организацией без нервных клеток; б, в — два этапа физиологической дифференцировки и появления отростков у клеток эктодермы; г — образование нервной сети, как у гидроидных стрекающих. Стрелками показано движение воды. 130 134 Нервные клетки концентрировались различными способами. Самым простым способом интеграции нервных сетей стало окологлоточное нервное кольцо. Его появление вполне оправдано тем, что оно находилось на границе поступления пищи в организм кишечнополостных. Пища была тем ведущим стимулом, который определял и оценивал успех морфологических изменений нервной системы. Тот, кто мог эффективнее контролировать поступление пищи, увеличивал свой метаболизм и репродуктивные возможности. Самым простым движением, позволяющим проиллюстрировать действие диффузной нервной системы, является реакция на механическое раздражение. Пресноводная гидра (Pelmatohydra oligactis)при малейшем раздражении сжимается в микроскопический комочек. Это происходит за счёт расположенных продольно в эктодерме и поперечно в энтодерме сократимых белков. Кроме генерализованной реакции, кишечнополостные могут дифференциально пользоваться отдельными щупальцами или их группами. Гидры способны передвигаться, чередуя при переворотах опору на подошву и ротовое отверстие. Тем не менее диффузная сеть с окологлоточным нервным кольцом была относительно медленно действующим устройством. Измеренная проводимость по нервной сети кишечнополостных составляет не более 5-20 см/с. Этого явно не хватает животным размером более 5 см, поэтому уже у актиний выделились участки нервной сети с высокой скоростью проведения (см. рис. II-4, в).В некоторых случаях она достигает 150 см/с, что делает актиний изощрёнными охотниками за значительно более эволюционно продвинутыми позвоночными. Окологлоточное нервное кольцо было явным достижением, но оно не могло дифференциально управлять всем телом или обеспечить контроль за свободным плаванием. Среди предков современных одиночных актиний явно были свободноплавающие существа. На это указывает двойная нервная сеть в их теле (см. рис. II-4, б). Одна диффузная сеть расположена под эктодермой в мезоглее и ничем не отличается от таковой у других кишечнополостных (см. рис. II-4, а).Другая нервная сеть лежит в той же мезоглее, но уже около энтодермы. Они связаны между собой только в зоне окологлоточного нервного кольца, которое начинает играть как интегрирующую, так и разделяющую роль. По-видимому, такие двойные сети возникли на заре эволюции нервной системы и были нужны для активного свободного плавания. Животное с автономной «эктодермальной» сетью могло активно двигаться в толще воды. Сокращение эктодермальных клеток позволяло животному перемещаться, не вовлекая в этот процесс пищеварительную систему. 135 Рис. II-4. Предполагаемые первые этапы (показано стрелками) усложнения строения нервной системы кишечнополостных с радиальной симметрией. а — однослойная сеть примитивных гидроидов; б — двойная нервная сеть актиний; в — нервная сеть актиний со скоростными проводящими цепями клеток; г — нервная сеть восьмилучевого полипа; д — нервный аппарат радиально-комиссурального типа. 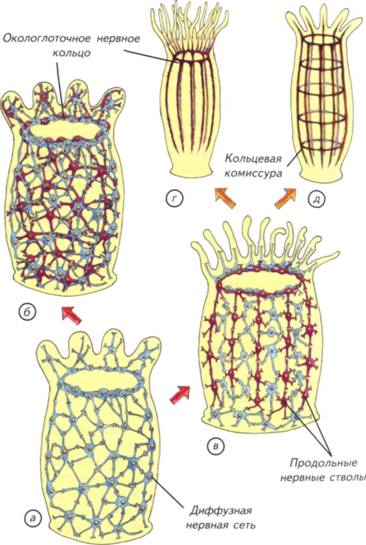 136 139 Рис. II-5. Общий вид и сечения основных вариантов строения нервной системы кишечнополостных и червей. а — нервная сеть восьмилучевого полипа; б — нервная система нематод; в — нервный аппарат радиально-комиссурального типа; г, e — нервная система турбеллярий; д — нервная система печёночного сосальщика. Синим цветом на сечениях обозначены нервные стволы. 140 147 Рис. II-7. Нервная система и ганглии различных членистоногих. а — чешуекрылые (Lepidoptera); б — двукрылые (Diptera); в — равноногие (Isopoda); г — веслоногие (Copepoda); д — ракушковые (Ostracoda); е-и — головной ганглий чёрного таракана; к, л — туловищный и подглоточный ганглии речного рака. Прямоугольником отмечен участок ганглия, показанный с большим увеличением. 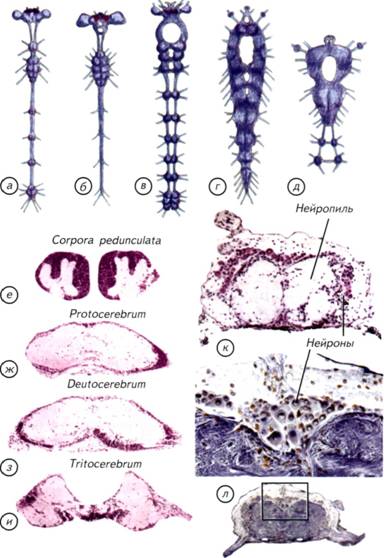 148 149 Рис. II-8. Нервная система различных беспозвоночных. а — первичнотрахейные (Onychophora); б — многоножки (Myriapoda); в — хелицеровые (Chelicerata); г — насекомые (Insecta). Процесс торсии — перекручивания внутренностного мешка и нервных стволов: до начала перекручивания (д) и после его окончания (е) у брюхоногих моллюсков (Gastropoda).  150 153 (Cephalopoda)моллюски со сложным инстинктивным поведением и способностью решать сложные экспериментальные задачи, не встречающиеся в естественных условиях обитания. § 23. Нервная система моллюсков Наибольший морфофункциональный контраст представляют собой организация нервной системы головоногих и двустворчатых моллюсков (рис. II-9; II-10, а).У двустворчатых моллюсков есть парные головные, висцеральные и педальные ганглии, соединённые комиссурами (см. рис. II-10, а). Нервные стволы, идущие от головных ганглиев, обычно замкнуты. Головные ганглии (cerebral ganglion) могут быть построены по различному типу. У мидий нейроны расположены в самом нейропиле ганглия (см. рис. II-9, и), а у сердцевидок и шаровок — по его периферии (см. рис. II-9, г). Различия в гистологическом строении ганглиев не сказываются на их функциях. Головные ганглии в обоих случаях иннервируют ротовые лопасти, передний аддуктор и мантию. Головные ганглии двустворчатых моллюсков составного типа. Они образовались в результате слияния церебрального и плеврального ганглиев. Висцеральный ганглий (visceral ganglion) также образовался в результате слияния с париетальным ганглием. Он обычно несколько меньше головного и расположен в каудальной части моллюска (см. рис. II-9, д).Висцеральный ганглий иннервирует мышцы ретрактора, заднего аддуктора, сердце, почку, сифон, половые органы и жабры. Пара педальных ганглиев (pedal ganglion) иннервирует мускулатуру ноги (см. рис. II-9, е). Они меньше других ганглиев, но имеют разветвлённые эффекторные отростки, оканчивающиеся среди мышц. Таким образом, нервная система двустворчатых моллюсков представлена всего тремя парами ганглиев, которые образовались в результате слияния. Это упрощение нервной системы связано с малоподвижным образом жизни и отсутствием рецепторных головных структур. Органы чувств у двустворчатых моллюсков достаточно специфичны и распределены по всему телу. На мантии, сифонах и жаберных нитях встречаются вторичные глаза, которые не связаны непосредственно с головным ганглием. Они имеют относительно сложное строение и могут воспринимать не только освещённость, но и направление её изменения. Это позволяет некоторым двустворчатым моллюскам перемещаться, пользуясь ногой или применяя реактивное движение — хлопая створками, как морской гребешок. У двустворчатых моллюсков хорошо развиты хеморецепторы — осфрадии и механорецепторные осязательные клетки. Есть и статоцисты, которые локализованы в крае ноги вблизи от педального ганглия. 154 155 Рис. II-9. Нервная система двустворчатых моллюсков (Bivalvia). а-е— С. galliпа corrugatula; ж-и— P. canaliculus. Красными точками на микрофотографии (в) показана локализация увеличенных ганглиев (г, д); г, и — головные ганглии (cerebralganglion); д — висцеральный ганглий (visceralganglion); e— педальный ганглий (pedalganglion). Стрелками показаны этапы препаровки моллюсков. 156 несколько вариантов торсии, что зависит от расположения жаберного аппарата или лёгких. Относительно скромная организация нервной системы брюхоногих моллюсков оказалась весьма эффективной как в воде, так и на суше. Большинство брюхоногих раздельнополы, хотя встречается и гермафродитизм с чередованием мужских и женских периодов размножения. В большинстве случаев гастроподы — подвижные животные, которые питаются растениями или хищничают. Однако встречаются и прикреплённые колониальные формы, такие, как каури. Подобные фильтраторы являются скорее исключением среди гастропод, чем правилом. Поведение брюхоногих не отличается интеллектуальными поступками и достаточно однообразно. Обычно брюхоногие питаются растительными остатками, непосредственно растениями или грибами. В этом случае их активность сводится к поиску пригодной пищи. Однако среди брюхоногих есть и настоящие хищники. Примером агрессивного хищника является тестацелла (Г. maugei), которая питается дождевыми червями. Она обладает достаточно развитыми органами чувств, чтобы охотиться на дождевых червей в их норах. Многие хищные улитки охотятся на себе подобных, но не брезгуют никакой животной пищей. В той или иной степени к хищничеству могут прибегать представители разнообразных семейств. Особую популярность в качестве объекта зоопсихологии приобрели головоногие моллюски (Cephalopoda).Осьминоги, кальмары и каракатицы являются, по мнению исследователей их поведения, своеобразными приматами мира беспозвоночных. Они ведут преимущественно активный образ жизни, перемещаясь по дну на многочисленных конечностях или плавают, используя принцип реактивного движения. Выталкивание воды из мантийной полости позволяет им быстро плавать в толще воды, но только задом наперёд. У головоногих в крайней форме проявляется цефализация нервных ганглиев (см. рис. II-10, в, ж, з).Ганглии с различными функциями сливаются над или под пищеварительной трубкой и образуют крупный головной ганглий. Поскольку головоногие могут достигать 20-метровой длины, размеры их головного ганглия могут быть сопоставимы с размерами мозга собаки. При таких размерах конструкции и традиционной для беспозвоночных ганглиозной организации нервной системы трудно представить себе эффективное метаболическое обеспечение. Именно размеры головных ганглиев головоногих неизбежно привели к возникновению внутримозговых полостей. У большинства головоногих моллюсков головные ганглии внутри полые. Однако надо оговориться, что речь идёт только о надглоточной части головного ганглия. 157 Рис. II-10. Нервная система двустворчатых, панцирных и головоногих моллюсков. Схема строения нервной системы двустворчатых моллюсков (а) и хитона (б); внешний вид нервной системы (в) и головного мозга (ж, з) кальмара; микрофотографии срезов головных ганглиев осьминога (Octopusvulgaris): r— брахиальный ганглий; д — висцеральный ганглий; e — церебральный ганглий. 158 Подглоточная часть обычно не имеет внутренней полости. Разделение головного ганглия на подглоточный и надглоточный отделы функционально вполне обосновано (см. рис. II-10, ж, з). Подглоточная часть головного ганглия состоит из брахиальных, педальных и висцеральных ганглиев, объединённых общим нейропилем (см. рис. II-10, г, д).Педальные ганглии уже внутри общей структуры разделены на ганглии щупалец и ганглии воронки. Брахиальные и педальные ганглии иннервируют щупальца и мантийную полость. Висцеральные ганглии отвечают за иннервацию внутренних органов и содержат центр автоматической регуляции дыхания. Полый внутри надглоточный ганглий является своеобразным интеллектуальным центром головоногих. В нём выделяют от 3 до 6 отделов. В самом сложном случае к ним относят буккальные ганглии (ротовые), три пары центральных и две пары церебральных ганглиев. Буккальные ганглии вынесены вперёд, не сливаются с головным ганглием и отвечают за иннервацию ротовой зоны передней части пищеварительной системы (см. рис. II-10, ж). Самыми ростральными надглоточными (уже головными) являются центральные ганглии, которые расположены поперёк оси головного центра. Их передняя пара контролирует механизм захвата, удержания, подведения к клюву, расчленения и заглатывания пищи; 2-я и 3-я пары центральных ганглиев хорошо развиты в связи с тем, что они отвечают за гидродинамическую систему плавания и контролируют обновление воды в воронке. Эти процессы осуществляются не непосредственно, а через контроль за исполнительными висцеральными центрами. Эти же центры контролируют перемещения головоногих при помощи щупалец, отвечают за осязательные ощущения и изменения направления движения при плавании. В центральных ганглиях находятся и центры управления окраской животных. Надо отметить, что головоногие могут очень широко изменять свою окраску из-за прямой иннервации пигментных клеток. Нервная система имеет непосредственные окончания на каждой пигментной клетке, что позволяет головоногим динамично изменять свой цвет при маскировке и смене настроения. Такой тип управления пигментацией позволил каракатицам использовать смену окраски тела как своеобразный цветовой язык. Во время брачных игр и оплодотворения самцы и самки ведут переговоры на языке смены цветовой гаммы покровов, поэтому не удивительно, что в тех же центральных ганглиях находятся и зрительные зоны. Туда приходит информация от глаз, которые у головоногих моллюсков самые большие среди животных. У крупных 159 162 беспанцирные, или бороздчатобрюхие моллюски (Aplacophora).Небольшая группа таких моллюсков напоминает скорее червей, чем моллюсков. Беспанцирные моллюски — морские животные длиной несколько сантиметров. Они ведут малоподвижный образ жизни и не обладают специализированными органами зрения. Их нервная система отдалённо напоминает нервную систему хитонов. Однако у беспанцирных моллюсков есть настоящие ганглии. Их нервная система состоит из окологлоточного нервного кольца и 4 продольных стволов, связанных нерегулярными комиссурами. Два брюшных ствола начинаются от подглоточных парных ганглиев, а боковые стволы — от асимметричного надглоточного. Нижние и верхние стволы заканчиваются в конце тела ганглиями, которые попарно соединены между собой. Надо отметить, что окологлоточное нервное кольцо у беспанцирных двойное, лежащее рострально, не связано с нервными стволами и имеет на брюшной поверхности парные небольшие ганглии. Под это описание вполне подходит нервная система и некоторых плоских червей, показанная на рис. II-5, г. Таким образом, беспозвоночные демонстрируют значительное разнообразие в организации нервной системы и поведении от примитивного поедания грунта до активной охоты на себе подобных и позвоночных. Хотя перечисленными группами далеко не исчерпывается многообразие мира беспозвоночных, в наши интересы входит только то направление эволюции, которое имеет отношение к проблемам возникновения хордовых. Мы рассмотрели только некоторые морфологические варианты строения радиальных нервных систем и их возможные преобразования в билатеральные конструкции. Попробуем подвести предварительный итог изложенным данным. |