Савельев. Происхождение мозга
 Скачать 1.82 Mb. Скачать 1.82 Mb.
|

|
§ 5. Энергетические расходы нервной системы Сопоставив размеры мозга и размеры тела животных, легко установить закономерность, по которой увеличение размеров тела чётко коррелирует с увеличением размеров мозга (см. табл. 1; табл. 3). Однако мозг является только частью нервной системы и не может рассматриваться отдельно от спинного мозга и периферической нервной системы. Масса одного мозга очень условно отражает энергетические затраты организма. Баланс между затратами на работу и приобретёнными результатами составляет суть работы нервной системы и затрагивает краеугольные принципы организации мозга, которые определили эволюцию нервной системы и стратегий поведения. Общий баланс энергетических затрат складывается из нескольких компонентов. Рассмотрим компоненты нервной системы, которые постоянно находятся в активном состоянии. К ним относятся все периферические отделы, поддерживающие тонус мускулатуры, контролирующие дыхание, пищеварение, кровообращение и т. д. Понятно, что отключение одной из таких систем приведёт к гибели организма. Нагрузка на эти системы постоянна, но не стабильна. Она меняется в зависимости от поведения. Если животное начинает питаться, то активность пищеварительной системы возрастает и расходы на содержание её нервного аппарата увеличиваются. Аналогично повышаются расходы на иннервацию и контроль за скелетной мускулатурой, если животное активно. Колебания этих энергозатрат относительно невелики, так как тонус мускулатуры или активность кишечника организм вынужден поддерживать и в состоянии покоя. Следовательно, работа периферических систем требует постоянных затрат, которые возрастают с повышением активности организма. Совершенно иная ситуация складывается с энергетическими затратами головного мозга. Головной мозг всегда активен независимо от состояния организма. Это связано с фундаментальными свойствами 41 47 Таблица 4. Отношение массы головного мозга к массе спинного мозга
механизмами принятия быстрых и адекватных решений. Активное регулируется более активным. Мозг должен работать с заметным опережением складывающейся ситуации, от этого зависят выживание и успех конкретного вида. Однако повышение метаболизма мозга приводит к неизбежному возрастанию затрат на его содержание. Возникает замкнутый круг: теплокровность требует усиления обмена, которое может быть достигнуто ещё большим повышением метаболизма нервной системы. Иначе говоря, успех теплокровных напрямую зависит от эффективности работы нервной системы. В реальной ситуации эта связь не столь прямолинейна, но общая тенденция сохраняется практически всегда. Для теплокровных животных размер тела становится критичным. Небольшие животные вынуждены постоянно решать основную, но не единственную задачу — искать пищу. Мелкие насекомоядные съедают ежедневно огромное количество пищи. Бурозубка ежедневно потребляет пищи в несколько раз больше массы собственного тела. В похожей ситуации находятся мелкие летучие мыши и птицы. У многих небольших животных возникли механизмы защиты организма от перерасхода энергии — торпидность. В этом состоянии колибри снижают метаболизм, частоту дыхания и температуру тела. На несколько часов животное впадает в своеобразную спячку, которая нужна для экономии энергии. Иначе говоря, мелкие теплокровные могут находиться в 48 49 (Thalarctos marinus)— 1/250, льва (Panthera leo)— 1/300, тапира (Tapirus americanus)— 1/500 и гиппопотама (Hippopotamusamphibius)— 1/1500. При увеличении массы тела доля нервной системы как отдельного органа существенно падает. Вместе с уменьшением относительных размеров нервной системы снижается и доля потребляемой ею энергии. В связи с этим крупное животное с большим мозгом находится в более благоприятном положении, чем небольшое. Для маленьких «головастиков» неизбежны переход на высококалорийную пищу и повышенный уровень метаболизма. Энергетические затраты на содержание мозга складываются из потребления кислорода, питательных веществ и поддержания водно-солевого баланса. § 6. Потребление мозгом кислорода Совершенно неверно связывать интенсивность метаболизма мозга с общим потреблением кислорода организмом (Шмидт-Ниельсен, 1982). Действительно, у землеройки потребление кислорода на 1 кг массы тела составляет 7,4 л/ч, а у слона — 0,07 л/ч. Однако это общее потребление кислорода, которое различается на порядки в разных частях тела как слона, так и бурозубки. Более того, у животных с разной биологией величина потребления кислорода одинаковыми органами тела также значительно различается. Представления о пропорциональном размерам тела изменении потребления кислорода мозгом остаются странным заблуждением. Если у любого млекопитающего потребление кислорода мозгом становится меньше 12,6 л/(кг-ч), наступает смерть. При таком уровне кислорода мозг может сохранять активность только 10-15 с. Через 30-120 с угасает рефлекторная активность, а спустя 5-6 мин начинается гибель нейронов. Иначе говоря, собственных ресурсов у нервной ткани практически нет. Ни у землеройки, ни тем более у слона не было бы никаких шансов выжить, если бы потребление кислорода мозгом не обеспечивалось специальными механизмами. Мозг получает кислород, воду с растворами электролитов и питательные вещества по законам, не имеющим никакого отношения к интенсивности метаболизма других органов. Величины потребления всех «расходных» компонентов относительно стабильны и не могут быть ниже определённого уровня, который обеспечивает функциональную активность мозга. Надо отметить, что мозг часто оказывает решающее влияние на метаболизм всего животного. Энергопотребление мозга не может быть ниже определённой величины. Обеспечение этого уровня достигается в разных систематических группах изменением скорости кровообращения в сосудах нервной системы. Причиной этих различий являются изменения числа капилляров в 1 ммз ткани мозга. Конечно, в разных 50 52 инстинктивен, а энергетические проблемы его содержания ставят непреодолимые барьеры для развития индивидуального поведения. На индивидуальном уровне может возникнуть только вариабельность применения врождённых программ поведения. Рис. I-8. Обменные процессы в головном мозге приматов. В метаболизме нервной системы можно выделить три основных динамических процесса: обмен кислорода и углекислого газа, потребление органических веществ и выделение продуктов катаболизма, обмен воды и растворов электролитов. Доля потребления этих веществ мозгом человека указана в нижней части. Обмен воды и растворов электролитов вычисляется как время прохождения всей воды организма через мозг. Верхняя строка — пассивное состояние, нижняя — напряжённая работа нервной системы.  53 54 Привилегированность нервной системы Нервная система многих животных (и особенно у млекопитающих) обладает одним свойством, которое ставит её в исключительное положение. Это свойство связано с её изолированностью от остального организма. Будучи основным механизмом интеграции работы внутренних органов и основой поведения, она является «инородным телом» для собственного организма. Иммунная система рассматривает нервную систему примерно как занозу. Если иммунная система «добирается» до мозга, то начинаются тяжёлые аутоиммунные процессы, малосовместимые с жизнью. Возникает парадоксальная ситуация. Нервная система потребляет огромную часть кислорода и питательных веществ всего организма, которую получает через кровь. Одновременно она должна быть тщательно изолирована от кровеносной системы, поскольку рассматривается клетками иммунной системы как инородный объект. С точки зрения биологической целесообразности видно явное противоречие. Основной интегрирующий орган не должен быть чужеродным для иммунной системы. Тем не менее это факт, которому довольно легко найти внятное объяснение. В головном мозге слишком много специализированных органических компонентов, которые больше нигде в организме не используются. Создавать в иммунной системе механизм их распознавания как «своих» клеток крайне сложно и неоправданно. Намного «дешевле» просто отделить нервную систему от всего остального организма. Этот принцип изоляции реализован в семенниках, яичниках и нервной системе. В самом общем виде изоляция нервной системы поддерживается при помощи гематоэнцефалического барьера, состоящего из нескольких типов специализированных клеток. Чтобы разобраться с изолированностью нервной системы от остального организма, надо рассмотреть элементарные принципы её строения. § 7. Гематоэнцефалический барьер Нервная ткань — это объединение специализированных клеток, которые воспринимают, обрабатывают, хранят и используют информацию о внешней среде и внутреннем состоянии организма. Этим функциям подчинено строение нервных клеток — нейронов. Нервные клетки имеют особенности, которые отличают их от других клеток организма (рис. I-9). Нейроны неодинаковы. Они различаются по размеру, форме ветвления дендритов и аксонов, выделению различных химических веществ и физиологической активности. Нейроны — 55 Рис. I-9. Строение нейронов и глиальных клеток. а — импрегнированные нейроны коры головного мозга человека. При такой окраске виден примерно 1 нейрон из 1000, что позволяет рассмотреть его отростки; б — глиальные клетки из мозга человека; в — строение нейрона и его аксона, закрытого глиальными клетками. 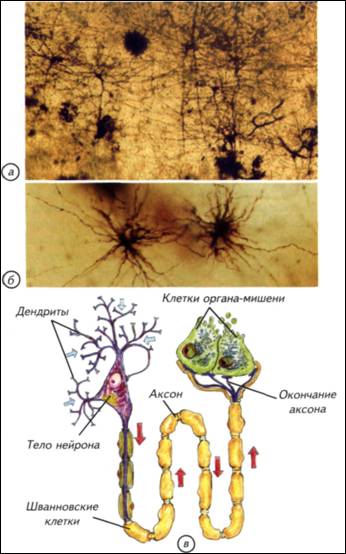 56 59 Рис. I-11. Срезы мозга и сосудистого сплетения (стрелки), расположенного в желудочках мозга различных позвоночных. Микрофотографии.  Спинномозговая жидкость фильтруется через эпендимные клетки нейрального происхождения. При низком кровотоке проницаемость стенок сосудистого сплетения невысока, но его площадь очень большая. У млекопитающих при высоком давлении крови сосудистое сплетение имеет крайне небольшие размеры. 60 62 Рис. I-12. Оболочки отростков нервных клеток (а, в)и синапсов (б). Электронные фотографии. Схема основных типов синаптических контактов нервных клеток (г). Синапсы и контакты увеличены. Оболочки отростков нервных клеток изолируют зоны проведения сигналов и увеличивают скорость их передачи. Синапсы обозначены зелёными стрелками. 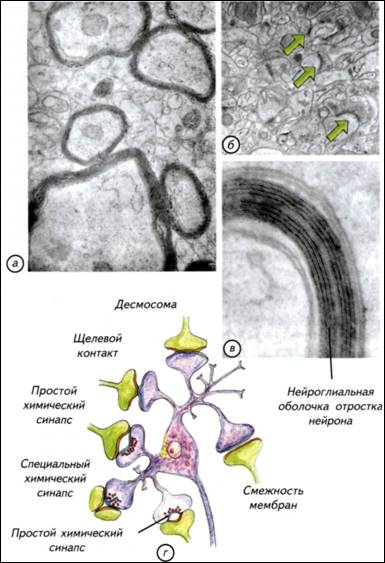 63 64 Взаимодействия между клетками Нервные клетки взаимодействуют между собой и с остальными тканями организма. Обычно это прямой контакт. Нервное окончание получает информацию или передаёт её клеткам органа, но это не обязательно. Нервные клетки могут синтезировать гормоны, нейропептиды или другие соединения. Они выделяются в кровеносное русло и распространяются по гуморальным законам. Гормоны используются как генерализованные носители информации для управления всем организмом. Иногда они специфичны только для определённого органа-мишени, но в целом гормональная регуляция очень неспецифична и определяет только общую тенденцию в поведении. Выброс половых гормонов происходит под влиянием нервной системы, но их присутствие в организме в конечном счёте подчиняет себе и работу мозга. Мозг «вызывает их к жизни» и сам подчиняется им. Так, в период гона у копытных стратегически меняется поведенческая активность. Половые гормоны оказывают столь заметное влияние на мозг, что все другие формы поведения отходят на второй план или становятся подчинёнными. Достаточно попробовать плоды блестящей дрессировки любимого домашнего пса в присутствии течной суки. В человеческом (приматном) сообществе действуют похожие законы. Весенняя гормональная активность преждевременно снимает шапки у мальчиков и оголяет коленки у девочек. Как правило, никакие «негормональные» доводы не действуют. Гормональная подчинённость нервной системы — это интеллектуальное горе человечества и гарантия его воспроизведения как биологического вида. Размножаться, драться и добывать пищу лучше с использованием гормональной поддержки организма. Древние викинги грызли край щита, доводя до нужного уровень адреналина перед боем. Словесная перепалка на кухне вызывает выброс мобилизирующих гормонов, а через 10 мин становится ясно, как много веских слов и аргументов ещё не высказано. Следовательно, гормональные межклеточные взаимодействия, запускаемые нервной системой, хороши, но инертны, неадаптивны и не поддаются динамическому контролю. Трудно представить, что, собираясь отчаянно спорить, человек будет колоть себя шилом для гормональной мобилизации. Ещё менее вероятен волк, грызущий свой хвост для охотничьего возбуждения. Для многих других видов гормональный контроль поведения позволяет просто статистически решать проблемы выживания. Для животных с выраженными генетическими программами поведения 65 69 процессы передачи сигналов происходят в нервной системе со скоростью, которая отражает динамику и продолжительность жизни организма. Скорость нервных процессов определяет активность животного и внутреннее восприятие времени. Надо отметить, что передаваемая информация кодируется частотой, последовательностью и продолжительностью активности импульсов, что обеспечивает высокую точность переносимой информации. Эти информационные сигналы различаются не только по принципам кодировки, но и по источникам — типам нервных клеток. Существует морфологическая и физиологическая классификация нейронов. По ведущим функциям нейроны физиологически подразделяются на сенсорные (афферентные), моторные, или двигательные (эфферентные), вставочные (ассоциативные, соединяющие афферентные и эфферентные) и нейросекреторные (гормональные клетки нервной системы). Эти типы не абсолютно жёсткие, а каждая клетка отчасти обладает всеми перечисленными функциями. § 9. Синаптические контакты нервных клеток Каждый нейрон способен воспринимать и передавать информацию. Он осуществляет это в специальных участках мембраны (см. рис. I-12). От тела нейрона обычно отходят одиночные аксоны, по которым нейрон передаёт потенциалы действия или градуальные сигналы. Кроме них, нейрон образует древовидные дендриты, которые являются основной зоной получения входящих сигналов. Однако и по дендритам нейрон может передавать сигналы другим клеткам. Как входящие, так и передаваемые нейроном сигналы проходят по специализированным участкам клеточной мембраны — синапсам. Количество синапсов существенно варьирует как у отдельных нейронов, так и у разных видов. Обычно синапсов бывает не менее 5000, а максимальное количество контактов может превышать 100 000. Синапсы различают по строению, типу передачи сигналов, используемому медиатору и физиологическому действию. В самом общем виде выделяют электрические и химические синапсы. Электрические синапсы считаются наиболее простыми и несущими линейную информацию. Они позволяют клеткам непосредственно влиять друг на друга без участия посредников или специальных механизмов. Передача сигнала происходит просто через щелевой контакт — специальное место в мембране (см. рис. I-12). В этой зоне белковые молекулы образуют специальные цилиндры, пересекающие межклеточное пространство и позволяющие сигналу переходить от клетки к клетке. Передача сигнала осуществляется очень быстро, но иногда только в одном направлении. Эти сигналы эффективны для согласования однозначных 70 72 Уровни организации нервной ткани Существует традиционное представление, что нервная система сложная или очень сложная. Однако сложная нервная система не столь недоступна для изучения, поскольку в её основе лежат те же принципы, что и в основе простой. Элементарным звеном нервной системы является нейрон, о котором уже говорилось. Нейрон — это специализированная клетка, которая способна получать, перерабатывать, хранить и передавать информацию. Однако нейрон, «вырванный» из своего окружения, не способен управлять поведением. Для создания хоть какого-нибудь поведения, отличающегося от физиологических реакций клеток растений, необходимо некоторое количество нейронов. Исследования простых нервных систем у круглых червей показали, что минимальная нервная система состоит из 30-100 нейронов. От такой сети уже можно ожидать реакций, напоминающих поведение более сложно организованных животных. Важно отметить, что даже при равном числе клеток существенные отличия в поведении возникают при особенностях морфологической компоновки нервной системы. Нейронам небезразлично, как они «организованы» и где «лежат» в организме. От этого зависит, как будет обрабатываться информация и насколько эффективно будет адаптироваться организм к изменяющимся условиям среды. § 10. Типы объединения нервных клеток Нервные клетки объединены в нервные системы различным образом. В простейшем случае эти элементы распределены вполне равномерно по всему телу животного или по большей его части (Anderson, 1990). Равномерное распределение нервных клеток обычно называют диффузной нервной системой. Самым известным животным с такой нервной системой является пресноводная гидра, которая может неспецифически реагировать на любое раздражение — сжиматься (см. рис. I-2). Её диффузная нервная сеть крайне проста по структуре и архаична по происхождению. По-видимому, это филогенетически самый древний способ объединения отдельных нервных клеток: клетки расположены равномерно по телу или органу животного и снабжены немногочисленными отростками, объединяющими нейроны в общую сеть (рис. I-13). Выделенных и протяжённых волокон в такой сети нет, а отростки клеток связывают только соседние нейроны. В сеть входят отростки, идущие от воспринимающих клеток эпителия или от рецепторов, а из сети отходят двигательные волокна, оканчивающиеся на поверхности мышечных клеток (см. рис. I-13). 73 Рис. I-13. Основные структурные уровни организации головного мозга, ганглиев и периферической нервной системы. Самый простой уровень — одиночная нервная клетка, которая может как рецептировать, так и генерировать сигналы. Такие клетки способны объединяться в системы с разделением функций (I). Более сложным вариантом являются ганглиозные скопления тел нервных клеток (II). Формирование ядер (III) или (слоистых) стратифицированных структур (IV) является наиболее сложным уровнем морфологической дифференцировки нервной системы. 75 Рис. I-14. Основные центры нервной системы позвоночных на примере лягушки. Головной мозг окрашен в красный цвет, а спинной — в синий. Вместе они составляют центральную нервную систему. Периферические ганглии — зелёные, головные нервы — оранжевые, а спинальные чувствительные ганглии — голубые. Взаимодействие центров осуществляется в результате постоянного обмена информацией между внешними и внутренними рецепторными системами. Обобщение и сравнение информации происходят в головном мозге. 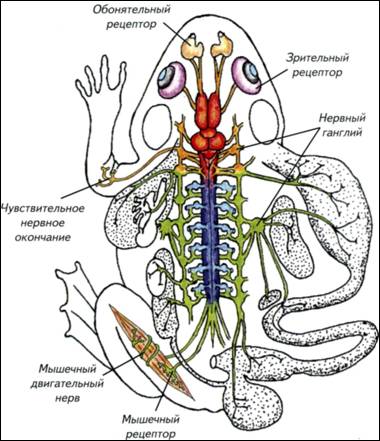 76 79 Рис. I-15. Гистологическое строение нервной системы позвоночных и беспозвоночных. Ганглии нервной системы имеют общий план строения как у позвоночных, так и у беспозвоночных животных. а — подглоточный ганглий речного рака; б — головные ганглии таракана; в — срез через головной мозг хорька; г — спинной мозг домовой мыши; д — ганглии дорсальных рогов спинного мозга мышонка. Срезы а, г, д окрашены по Маллори, б, в — по Нисслю. 81 Рис I-16. Основные тенденции усложнения структурной организации нервной системы. Синими стрелками обозначен условный этап появления нервных клеток, красными — основной путь церебрализации беспозвоночных, а зелёными — позвоночных животных. В обоих случаях наиболее сложная нервная система организована по ганглиозно-ядерно-корковому типу. Основными отличиями позвоночных являются больший размер мозга, наличие мозговых желудочков и замкнутое кровообращение. 86 Органы чувств и эффекторные системы Нервная система возникла как способ быстрой адаптации организма к изменению внешней, а затем и внутренней среды, поэтому источники информации об этих изменениях эволюционировали вместе с мозгом. Первоначально, как указывалось ранее, клеточные предшественники нервной системы были однотипны. Они отличались от окружающих клеток только способностью быстрее проводить сигналы, что и выделило их в самостоятельную и быстродействующую нервную систему. Однако реакции на внешние раздражители становятся совершеннее при точном распознавании источника воздействия. Чем лучше идентифицирован раздражитель, тем адекватнее будет ответ организма. Можно сформулировать основные требования к распознаванию сигналов, которые лежат в основе возникновения и эволюции органов чувств. 1. Различение типа и природы воздействия. Органы чувств должны определять природу воздействия. Она может быть химической, механической и электромагнитной. Этими свойствами в определённой степени обладают любые живые клетки. 2. Величина воздействия. Рецепторный аппарат должен определять величину воздействия. Чем шире диапазон чувствительности, тем точнее будет реакция организма. 3. Получение контактной и дистантной информации. Если понадобится, органы чувств должны получать информацию на расстоянии от предмета. Чем больше дистанция эффективной работы сенсорной системы, тем больше шансов у животного подготовиться к реальному контакту с объектом или избежать его. 4. Работа внешних органов чувств должна постоянно сопоставляться с работой внутренних систем организма. Для такого сравнения необходимы специальные центры, где можно было бы сравнить информацию из внешней и внутренней среды. Иначе говоря, внутренние рецепторные системы оказывают не меньшее, а часто и большее влияние на выбор конкретной формы поведения. На заре возникновения многоклеточных организмов одинаковые клетки первичной нервной системы не различались по возможностям. Они могли воспринимать одни и те же сигналы с одинаковой интенсивностью. При повышении требований к точности рецепции проблема может решаться двумя путями. В самом простом случае это увеличение количества и повышение чувствительности универсальных рецепторных клеток нервной системы. При этом никаких принципиальных изменений 87 89 Рис. I-17. Примеры строения некоторых дистантных и контактных рецепторов нервной системы позвоночных. Непосредственно рецептировать и передавать сигналы в нервную систему могут только первичночувствующие клетки (обонятельные клетки и свободные нервные окончания). Как правило, эти функции разделены между разными типами нервных клеток. Красным цветом отмечены рецептирующие клетки, зелёным и синим — передающие нейроны. Высокая чувствительность часто связана с модуляцией сигнала, проходящего через рецепторную клетку. Волосковые клетки получают входной сигнал по эфферентному волокну (голубая клетка), а его считывание происходит при помощи афферентной клетки, которая в ганглии сравнивает разницу между посланным и возвратившимся сигналами. 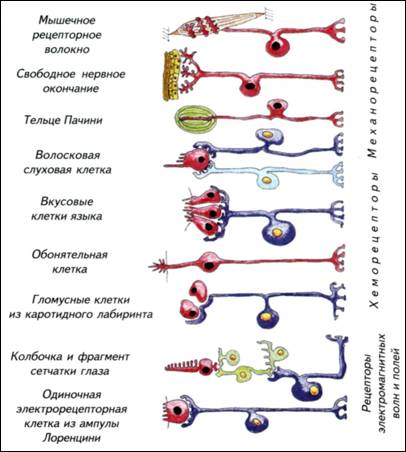 90 91 Рис. I-18. Гистологические срезы через тело личинки (головастика) лягушки и различные органы чувств. а — горизонтальный срез через тело головастика (показано в верхнем правом углу); б — срез через обонятельный мешок; в — срез через сетчатку и хрусталик глаза; г — поперечный срез через участок внутреннего уха. 92 Описанным способом работают фоторецепторные клетки глаза и пинеального комплекса гипоталамуса. Однако существует несколько специализированных электрорецепторов, которые воспринимают электромагнитные волны не в световом диапазоне. К ним относят этмоидные органы термочувствительности некоторых змей и ампулярные электрорецепторные органы рыб. Инфракрасный диапазон используется вторичночувствующими клетками змей для нахождения теплокровных жертв. Ампулами Лоренцини рыбы рецептируют изменения напряжённости электромагнитного поля, которое создают другие организмы. Электро-рецепция известна не только у пластиножаберных и костных рыб. Среди амниот такой способностью наделены некоторые птицы и утконос. Интересно, что рецептируемая напряжённость поля одинакова у рыб и утконоса. Однако поведенческая реакция проявляется у утконоса при раздражении в 10 раз слабее, чем у рыб. Существуют косвенные свидетельства возможности рецепции дельфинами электромагнитного поля Земли. Эту способность связывают с присутствием в нейронах и мозговых оболочках ряда органических соединений, которые могут менять ориентацию даже в слабых электромагнитных полях. Следует подчеркнуть, что в самой центральной нервной системе нет никаких специализированных рецепторов, хотя клетки многих отделов мозга реагируют на изменение концентрации гормонов в крови. Прямое оперативное вмешательство на мозге не вызывает болевых ощущений. Несколько иная ситуация с парасимпатическими ганглиями, расположенными в непосредственной близости от внутренних органов. За последние годы накопилось немало свидетельств о существовании специализированных свободных нервных окончаний, иннервирующих нейроны, находящиеся в периферических парасимпатических ганглиях. Несмотря на глубокие различия между позвоночными и беспозвоночными животными, мозговое представительство органов чувств построено по общему принципу (рис. I-19). Каждый специализированный орган чувств представлен в головном мозге скоплением клеток, которые обрабатывают поступающие сигналы и сравнивают их с активностью других органов чувств. Чем сложнее рецепторная система и чем большую роль в жизни животного она играет, тем морфологически выраженнее представительство данного органа чувств в головном мозге. Морфологическая структура мозга как позвоночных, так и беспозвоночных является своеобразным зеркалом их способности воспринимать мир. Однако никакой поведенческой адаптации животных к внешней среде без развитой и быстро работающей эффекторной системы 93 Рис. I-19. Организация головного мозга позвоночного (акулы) и головных ганглиев беспозвоночного (насекомые). Сходные анализаторные системы окрашены в идентичные цвета. а — позвоночное; б — насекомое. Несмотря на глубокие морфологические различия в эволюционных стратегиях становления головных отделов нервной системы, обе конструкции имеют множество общих черт. Обработка информации от анализаторов разделена в специализированных структурах головных ганглиев и головного мозга, а ассоциативные системы обособлены и часто стратифицированы. 94 представить невозможно. Если бы животные и человек могли воздействовать на окружающий мир «силой мысли», то в мясистой филейной части не было бы нужды. Даже самая великолепная мысль не может вырваться из мозга самостоятельно. Она доходит до человечества только через сокращение мускулатуры языка и конечностей. В связи с этим эффекторный аппарат управления организмом следует рассмотреть несколько подробнее. § 14. Эффекторные системы В основе эффекторных систем лежит всё та же способность клеток воспринимать сигналы химической, механической и электромагнитной природы. Однако источником этих сигналов становится не внешняя среда, а сама нервная система. Внутренние органы и скелетная мускулатура получают управляющие сигналы от нервной системы или её посредников. Изменение состояния этих эффекторных систем приводит к физической модификации наблюдаемого поведения. Надо отметить, что у животных и человека нет переизбытка вариантов воздействия на внешнюю среду. Эффекторные системы включают в себя несколько компонентов. Основным реальным способом воздействия на внешний мир является механическая активность животного. Этот непосредственный эффекторный механизм построен на различных вариантах деформации тела. Тело можно перемещать в пространстве по гидродинамическим законам, как это делают кальмары, выбрасывая порции воды из-под мантии. Неплохой вариант аппарата движения представляют гидростатические амбулакральные ножки, которые используют морские звёзды. Однако основной способ двигательной активности у животных связан с сокращением мускулатуры. Скелетная мускулатура состоит из нескольких типов поперечнополосатых мышц, которые различаются по величине, продолжительности усилий и типу метаболизма. Поперечная исчерченность этих мышц обусловлена упорядоченным расположением слоёв актиновых и миозиновых филаментов. Филаменты способны к движению относительно друг друга с затратой энергии, что и является основой механической активности мышц. Сходное строение имеет и сердечная мышца, но между её клетками расположены электрические соединения. Они позволяют распространяться электрическому току от клетки к клетке. В скелетной мускулатуре обычно выделяют тонические, медленные и быстрые фазические волокна. Тонические мышечные волокна не могут совершать одиночных сокращений и сокращаются медленно. Их сокращение полностью контролируется нервными импульсами мотонейронов. Эти мышцы распространены у амфибий, 95 97 Рис. I-20. Эффекторные системы организма позвоночных. а — эффекторные нервные волокна управляют работой клеток гладкой и поперечнополосатой мускулатуры, наружных секреторных систем и желёз внутренней секреции; б — в покровах многих первичноводных позвоночных расположены пигментные клетки, которые меняют окраску животного в зависимости от окружающей среды. Эти клетки имеют центральную нервную регуляцию, а управляющий центр связан со зрительной системой животного. 98 Память и забывание Природа памяти остаётся одной из самых излюбленных тем для «физиологических» фантазий. Тема столь популярная и доступная для натурфилософских и умозрительных рассуждений, что мимо неё не прошёл ни один популярный мыслитель. Надо отметить, что и подходы к решению проблемы памяти были столь же общественно популярны, сколь далеки от реальности. В соответствии с модой времени предлагались нематериальные формы памяти, которые прекрасно агрегировались с душой. При этом память легко заселялась в тело или «удирала» из него в зависимости от теологической оценки поведения человека. Начиная с XIX в. память пытались объяснять более или менее реальными химическими или физиологическими процессами. До настоящего времени распространены биохимические модели запоминания. Они особенно расцвели после открытия структуры ДНК в эпоху начала молекулярно-биохимических исследований. Дело дошло 100 до того, что были предложены экспериментальные «доказательства» существования «белковых молекул памяти». На них, как на популярную в те времена аудиоленту, записывалась некая информация о событиях. Затем она прокручивалась, расшифровывалась и проявлялась в сигналах нейронов. Следовательно, достаточно ввести необученным животным «белки памяти» от обученных, как они начнут решать сложные задачи, что, собственно, и было сделано. Прошедших обучение планарий («отличников») разрезали на кусочки и скармливали ничего не ведавшим собратьям. «Неучёных» каннибалов запустили в незнакомый лабиринт, где они проявили сноровку своего учёного обеда. Этот эксперимент стал широко известен, так как полностью соответствовал страстному увлечению биохимической природой памяти. Понятно, что недостоверность результатов была быстро установлена. Попытка приёма внутрь «информационных» молекул, которые будут разрушены пищеварительными ферментами, выглядит, как анекдот. Тем не менее подобные странноватые эксперименты регулярно обсуждаются до настоящего времени. В неявной форме туманные рассуждения о РНК-глиальных взаимодействиях как хранилище памяти неких информационных нейропептидов, соматических носителях памяти и особых информационных колебаниях регулярно возникают в литературе. Трудно не согласиться с тем, что пассивное (энергонезатратное) хранение любых записей на любом носителе дало бы мозгу огромные преимущества. Однако эти фантазии перечёркиваются печальным опытом клинической смерти человека. Примерно через 6 мин после начала клинической смерти из мозга начинают необратимо исчезать воспоминания, а через четверть часа о личных воспоминаниях говорить уже не приходится. Если бы память хранилась на каком-либо энергетически независимом носителе, то она бы могла восстановиться. Этого не происходит, что означает динамичность памяти и постоянные энергетические затраты на её поддержание. Проблемы механизмов памяти трактуют как некий круг широких и разнонаправленных процессов мозга, который понять, по-видимому, не дано. Столь приятный вывод стимулирует деятельность мозга и позволяет перечислить безусловный набор фактов, которые отражают существование или исчезновение памяти. § 15. Механизмы памяти 1. Память — это функция нервных клеток. При синдроме Корсакова, рассеянном склерозе, ишемической болезни мозга, когда дегенерируют нейроны, память исчезает. 101 10З Рис. I-21. Запоминание и забывание информации. Кратковременная память образуется на основании уже имеющихся связей. Её появление обозначено оранжевыми стрелками на фрагменте б. По одним и тем же путям циркулируют сигналы, содержащие как старую (фиолетовые стрелки), так и новую (оранжевые стрелки) информацию. Это приводит к крайне затратному и кратковременному хранению новой информации на базе старых связей. Если она не важна, то энергетические затраты на её поддержание снижаются и происходит забывание. При хранении «кратковременной», но ставшей нужной информации образуются новые физические связи между клетками по фрагментам а-б-в. Это приводит к долговременному запоминанию на основании использования вновь возникших связей (жёлтые стрелки). Если информация долго остаётся невостребованной, то она вытесняется другой информацией. При этом связи могут прерываться и происходит забывание по фрагментам в-б-а или в-а (голубые стрелки). 104 109 во вполне материальном виде. Забывание — биологически очень выгодный процесс. При исчезновении любого самого короткого информационного цикла происходит экономия на синтезе АТФ, медиаторов, мембран и на аксонном транспорте. Всё, что приводит к уменьшению энергетических расходов мозга, воспринимается как биологический успех. Мозг не «догадывается» об информационной ценности памяти. Он стремится экономить на её хранении. Ему безразлично, на что затрачивается энергия, главное — её количество, а не качество. Мозг стремится заставить весь организм экономить и делать стратегические запасы. Для мозга человека биологическое прошлое важнее любых социальных законов. Сколько ни стойте перед холодильником с разносолами, мозг никогда не «поверит», что завтра будет столько же еды. Он заставит организм набить желудок с большим запасом. С тем же обезьяньим упорством мозг человека стремится не расходовать энергию на затратное поддержание любой информации. Он с одинаковым удовольствием забудет и номер банковского счёта, и рекламный листок из почтового ящика. В связи с этим забывание любой информации происходит намного легче и приятнее, чем её запоминание. Мышление Исторически сложилось так, что мышление представляет собой почти философский вопрос. Если о механизмах памяти существуют хотя бы странноватые гипотезы, то о мышлении нет даже таких. О занятных умозрительных психолого-лингвистических фантазиях Юма, Милля и Спенсера уже достаточно сказано, чтобы не заниматься историей науки. Основная проблема столь незатейливого процесса, как мышление, связана с самими мыслителями, а не с существом дела. Стандартный подход исследователей мышления сводится к объяснению привычными понятиями свежевыдуманных и менее привычных терминов. Туманные натурфилософские объяснения биологического смысла психологических понятий ещё более усиливают сомнения в компетенции упомянутых сочинителей. Суть надуманной философской проблемы мышления оформлена в виде неразрешимого «гносеологического вопроса». Неразрешимость достигнута противопоставлением двух теорий происхождения и организации мышления. По «рационалистической» теории, законы мышления абсолютно априорны. Они предшествуют опыту и являются базой для его осуществления. Мышление наследуется, его принципы неизменны. С другой, «эмпирической», точки зрения, законы мышления апостериорны. Все они вытекают из опыта и являются благоприобретёнными свойствами мозга. На самом 110 113 Рис. I-22. Головной мозг основных представителей позвоночных с латеральной поверхности. Основные отделы мозга обозначены одинаковым цветом. Красный цвет и крестообразная штриховка показывают расположение ассоциативных зон мозга. 114 мозга, хотя роль зрительных центров среднего мозга остаётся значительной. Только у млекопитающих ассоциативные центры сосредоточены в развитом неокортексе переднего мозга. Это стало возможно благодаря увеличению размеров мозга в целом и переднего мозга в частности. Избыток клеток в анализаторных и ассоциативных зонах дал огромные преимущества животным и существенно усложнил мышление. § 18. Биологические проблемы мышления Обладатели крупного мозга становятся своеобразными заложниками его размеров. Нравится или не нравится животному, но в большом количестве нейрональных связей постоянно фиксируется повседневная информация о внешнем мире и внутреннем состоянии (рис. I-23). Чем больше мозг, тем разнообразнее информация и дольше она в нём хранится. Казалось бы, львы в стабильном прайде не отличаются высокой активностью. При самой пассивной форме поведения и увеличении продолжительности сна до 2/3 сут мозг всё равно продолжает активно работать. Он накапливает информацию и непрерывно образует новые связи между нейронами. Нейроны коры каждые 2-3 дня формируют новый синапс, а один раз в 1,5 мес — небольшую коллатераль. При этом примерно столько же старых связей разрушается. Иначе говоря, мозг непрерывно изменяет материальную базу нашего мышления. Даже полное бездействие в конце концов приведёт к формированию случайного набора нервных связей, который вызовет неожиданное и совершенно «немотивированное» поведение. На самом деле «мотивация» была, но она не связана с неким определённым индивидуальным опытом или конкретным событием. Побуждение к действию возникло из-за вновь образованных межнейронных связей. Они случайно соединили разнообразные хранилища образов, слуховых стимулов, запахов и моторных навыков. Появилась доныне не существовавшая связь между явлениями, что побудило животное или человека к формально «немотивированному» действию. Мозг с огромным количеством морфогенетически активных нейронов непрерывно создаёт новые связи. Он неизбежно будет накапливать и утрачивать различные сведения, а поведение станет непроизвольно меняться. Хранение информации автоматически приводит к её сравнению со следующей порцией аналогичных сведений. Даже ежедневная информация об одном и том же пастбище и результатах поглощения травы различается у каждой конкретной коровы. Элементарное сравнение приводит её на поле с молодыми всходами, а не в сосновый лес. 115 Рис. I-23. Структурные механизмы принятия решения. Допустим, что в памяти существуют три явления (стрелки), которые представлены в виде четырёхклеточных нейронных комплексов: а-в. В них хранится информация от трёх органов чувств: зрения, обоняния и вкуса. Каждый тип информации обозначен собственным цветом стрелок. Мозг пытается связать разнородную информацию, что приводит к перераспределению потока крови и образованию новых связей. Формируются связи внутри и между комплексами. Они позволяют сигналам одного комплекса переходить в поле другого и «сравниваться». Идентичность движения сигнала рассматривается как сходство явлений, что позволяет сопоставить активности различных органов чувств и связать события между собой. Это решение первичной задачи сравнения, а новые пути дают вариабельность ответов. На основании интеграции движения сигналов возникает совершенно новая система связей (красные стрелки), которая является решением проблемы, известной как инсайт, или озарение — неожиданное решение возникшей проблемы. 116 |