Савельев. Происхождение мозга
 Скачать 1.82 Mb. Скачать 1.82 Mb.
|

Глава III. Становление мозга амниот § 34. Репродуктивные стратегии амниот К амниотам относят рептилий, птиц и млекопитающих. Группа объединена по эмбриологическому принципу устройства оболочек зародышей. Они принципиально отличаются от неклеточных икряных мешков или стенок икринок амфибий (Ромер, Парсонс, 1992). У амфибий размножение обычно связано с водой, а внезародышевые оболочки отсутствуют. Икринки откладываются самками амфибий в воду, где они оплодотворяются сперматозоидами самцов. В икринках развиваются эмбрионы, которые трансформируются в личинки и переходят к самостоятельному питанию в воде (рис. III-1). Затем следует метаморфоз с выходом на сушу или половое созревание для водных и неотенических форм амфибий. Оплодотворение созревших яйцеклеток в воде является древнейшим способом размножения, характерным для первичноводных позвоночных. У многих амфибий происходит внутреннее оплодотворение, хотя эмбрионы и личинки по-прежнему развиваются в воде (см. рис. III-1; рис. III-2). По-видимому, у лабиринтовых предков амниот размножение отличалось от архаичных рыб (Dodd, Dodd, 1985). Первоначально они могли иметь наружное оплодотворение и откладывать икру в укромных местах своих охотничьих лабиринтов. Однако риск уничтожения потомства в таких условиях очень велик. Снизить гибель потомства можно было только одним способом — носить икру с собой. Появление такой заботы о потомстве довольно обычно у костистых рыб и неоднократно происходило в процессе эволюции (Jameson, 1988). Забота о потомстве должна была неизбежно привести к появлению внутреннего оплодотворения с последующим вынашиванием развивающейся икры внутри тела (см. рис. III-1; III-2). Переход к живорождению стал у лабиринтовых амфибий принципиальным шагом на пути адаптации к наземной среде. Размножение стало отчасти независимым от водной среды. У архаичных амфибий существовали все возможные варианты наружного и внутреннего оплодотворения, длительного или краткосрочного вынашивания развивающейся икры (см. рис. III-2, а-г).Наиболее успешными стали животные с длительным вынашиванием зародышей. Судя по всему, они могли появляться на свет уже полностью готовыми к самостоятельному существованию. При откладке икры в воду зародыш может иметь минимальный запас желтка (см. рис. III, а). Личинка рано 252 выходит из икринки и начинает питаться самостоятельно. При удлинении срока развития потребность в желтке или другом источнике питательных веществ резко возрастает. Эта проблема решается несколькими способами. Можно использовать индивидуальный запас желтка или богатые секретом яйцеводы материнского организма. Зародыши некоторых живородящих пластиножаберных питаются стенкой яйцеводов вплоть до выхода из материнского организма. Однако чаще всего просто увеличивается индивидуальный запас желтка, в результате зародыш может долго находиться в организме матери. Он появляется на свет уже сформированным организмом, готовым к самостоятельному питанию. Такие варианты заботы о потомстве часто встречаются во всех группах первичноводных позвоночных. Ни к каким особым эволюционным достижениям такое внутреннее развитие, казалось бы, привести не должно. Тем не менее одна особенность биологии отличает древних амфибий от первичноводных позвоночных. Амфибийный образ жизни предусматривает пребывание на суше, где механические деформации тела под действием гравитации заметнее, чем в водной среде. Механические воздействия на репродуктивные органы, ещё не защищённые специализированными элементами скелета, неизбежно должны были привести к развитию компенсаторных демпферов — дополнительных оболочек (см. рис. III-1, б). Если бы зародыш находился вне организма матери, то никаких специальных внезародышевых оболочек не понадобилось, но на суше деформационные воздействия на органогенез внутри яйцеводов весьма велики. Это связано с тем, что на ранних стадиях развития позиционная информация в морфогенетически активных зачатках эмбриона кодируется при помощи механогенетических процессов. Суть этого явления состоит в изменении проницаемости механозависимых ионных каналов при формообразовании. Они обеспечивают каждой клетке индивидуализированный позиционный сигнал, который биомеханически интегрирован со всем зачатком или растущим органом. Благодаря этим взаимодействиям клетка периодически получает управляющий позиционный сигнал и запускает региональные гистогенетические процессы (Савельев, 1993). Надо отметить, что этот механизм регуляции раннего эмбрионального формообразования характерен для всех позвоночных и предопределяет необходимость биомеханической защищённости развития. Понятно, что столь ненадёжный механизм весьма уязвим. Даже при незначительных статичных деформациях зародыша легко возникают множественные аномалии развития и эмбрион погибает. Любой переход к 253 Рис. III-1. Преобразования внезародышевых оболочек (показано фиолетовыми стрелками) при переходе к наземному образу жизни. а — неклеточные оболочки икры у рыб и амфибий; б — развитие в организме матери — увеличен желточный мешок, складки амниона; в — появление аллантоиса для накопления продуктов распада; г — использование аллантоиса для обмена веществ с материнским организмом; д — схема яйца рептилий и птиц в яйцевой оболочке. 254 255 Рис. III-2. Возможные пути преобразования эмбрионального развития при переходе к наземному образу жизни (показано большими стрелками). Первичным является наружное оплодотворение и внешнее развитие икры (а). Затем возникло внутреннее оплодотворение и развитие (б), которое у амфибий вернулось к исходному варианту (в). Живорождение при наземном образе жизни (г) первично по отношению к яйцекладущим и плацентарным животным (д, е).  256 261 Рис. III-3. Появление первых амниот привело к невероятному разнообразию жизненных форм. За относительно непродолжительный период были освоены суша, воздушная и водная среда обитания. Такой прогресс смогли обеспечить совершенный набор физиологических адаптаций и головной мозг с новыми принципами организации. Контуры тел исчезнувших и современных рептилий дают представление о видовом разнообразии этой группы. 262 Seismosaurus).При размерах тела более 40 м и массе около 100 т эти животные обладали головным мозгом не больше мозга современной крупной собаки. Основные физиологические проблемы этих гигантов решались не головным, а спинным мозгом, на уровне автономной нервной системы. Приблизительные оценки массы спинного мозга показывают, что его размеры и масса намного превышали аналогичные показатели головного мозга. По-видимому, у крупных (более 5 м) рептилий головной мозг не превышает 1/10-1/15 массы спинного мозга. Примерно треть массы спинного мозга приходится на плечевое и поясничное утолщения, которые хорошо развиты у современных рептилий (рис. III-4). Вполне допустимо предположить, что у большинства динозавров спинные и поясничные утолщения имели терминальные желудочки, как у современных птиц и медведей. Это позволяло эффективнее осуществлять метаболизм в наиболее функционально активных участках спинного мозга. Однако спинной мозг является исполнительной частью нервной системы. Он только осуществляет набор автономных программ, но не принимает решения. Вся поведенческая активность контролируется головным мозгом. У архаичных рептилий он имел невероятно скромные размеры, но позволял им находить пищу, размножаться и конкурировать с другими видами. Проводя глобальные аналогии, можно сказать, что относительная масса мозга динозавров была намного меньше, чем у современных насекомых. То же заключение можно сделать и в отношении амфибий. Однако они не смогли достигнуть уровня расцвета рептилий. Это позволяет сделать вывод о принципиальном эволюционном изменении в конструкции мозга при появлении древних амниот. Таким образом, разнообразие форм исчезнувших и современных рептилий практически не имеет аналогов в истории позвоночных. На этом фоне однотипность строения их мозга выглядит парадоксально. Принципы его конструкции практически не изменялись как у архаичных, так и у современных рептилий. Анатомические различия внешнего строения мозга в основном связаны с количественными, а не качественными изменениями. В соответствии с адаптацией к конкретной среде обитания может преобладать тот или другой отдел мозга, но принципы его строения сохраняются неизменными, а количественное (макроанатомическое) представительство анализаторов или моторных центров варьирует в широких пределах. Эти изменения меньше отражаются на внешней форме мозга, чем у первичноводных позвоночных (см. рис. III-4). Стабильность конструкции головного мозга рептилий говорит о ряде 263 Рис. III-4. Внешний вид головного, спинного мозга и плечевого нервного сплетения рептилий на примере современных диапсид. а, б — аллигатор; в — варан. Оба вида принадлежат к различным группам диапсид: аллигатор относится к архозаврам, а варан к лепидозаврам. Конструкция их головного и спинного мозга принципиально не отличается от «рептилийного» архетипа. 264 принципиальных приобретений, которые оказались настолько универсальны, что не нуждались в качественной перестройке миллионы лет. Новые по сравнению с амфибиями свойства мозга рептилий дали этой группе значительные эволюционные (поведенческие) преимущества, позволившие освоить всю сушу, воздушную и водную среду. § 36. Общий план строения нервной системы рептилий При масштабном освоении суши мозг рептилий изменился. Однако его морфологические перестройки, на первый взгляд, не были столь принципиальными, как у амфибий. Поверхностный анатомический анализ показывает, что древний «амфибийный» архетип строения центральной нервной системы не подвергся радикальным изменениям (см. рис. III-4; рис. III-5; III-6). Этот поверхностный взгляд на морфологию нервных центров рептилий может ввести в заблуждение. Только рептилии смогли эффективно освоить сушу, доминировать в воде, подняться в воздух и стать первой доминирующей группой позвоночных. Основой для такой широкой и полиморфной радиации был не только бесконечный пищевой ресурс, но и эффективная система управления организмом — мозг. При появлении рептилий произошёл ряд существенных перестроек нервной системы, которые затем стали базовыми для всех амниот. По-видимому, эти изменения были схожими, хотя и могли возникать совершенно независимо. Основная трудность состоит в том, что первичный архетип строения нервной системы был замаскирован дальнейшими адаптивными специализациями. Его реконструкция возможна только при выделении ключевых и повторяющихся структурных элементов в организации мозга рептилий. К сожалению, современные рептилии скромно представлены довольно специализированными видами. Это несколько затрудняет поиск древних конструкций мозга, принадлежавших их далёким предкам. Рассмотрим общий план строения нервной системы рептилий. Их нервная система развита значительно лучше, чем у амфибий и первичноводных позвоночных. Спинной мозг имеет цилиндрическую форму с плечевым и поясничным утолщениями, соответствующими поясам конечностей. Сзади спинной мозг продолжается, постепенно сужаясь, в начало хвостового отдела позвоночника. Головной мозг более широк и массивен, чем у амфибий, особенно у крокодилов и черепах. Для рептилий характерны изгибание ствола и стремление крупных полушарий надвинуться на лежащие позади промежуточный и средний мозг (см. рис. III-6; рис. III-7; III-8). Полушария переднего мозга без резких границ переходят в обонятельные луковицы, от которых начинаются обонятельные нервы. 265 Рис. III-5. Голова геккона (Gekko gekko)с вписанным в неё головным мозгом и гистологические срезы через основные отделы. а — полушария переднего мозга; б — промежуточный мозг; в — средний мозг; г — задний мозг; д — каудальный конец продолговатого мозга; e — спинной мозг. Срезы соответствуют уровням расположения и обозначениям сечений на схеме. 266 267 Рис. III-6. Головной мозг современных рептилий различных систематических групп и схема мозга анамний и холоднокровных амниот. а — хамелеон (Chamaeleolateralis); б — красноухая черепаха (Pseudemysscripta); в — аллигатор (Alligatormississippiensis); r — сетчатый питон (Pythonreticulatus); д — схематическое изображение мозга рептилий; e — схема мозга амфибий. Голубые и зелёные стрелки показывают связи между отделами мозга. Грибовидные расширения над двойной полосой обозначают ассоциативный центр, а под полосой — адаптивное увеличение размеров отдела. Красные стрелки показывают связи ассоциативного отдела с другими отделами мозга. 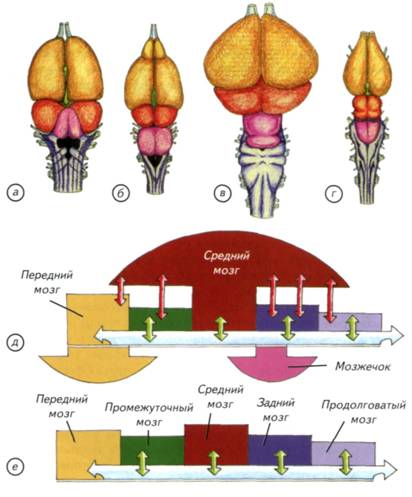 268 к 269 Рис. III-7. Гистологическое строение рептилий и ассоциативных центров головного мозга. а — фронтальный срез через полушарие переднего мозга с основными ассоциативными кортикальными структурами, расположенными между маркёрами границ; б — срез через многослойную крышу среднего мозга геккона; сагиттальный срез через теменной глаз (в) и лабиринт (г) Lacerta agilis; д — сагиттальный срез через эмбрион L agilis перед вылуплением. Объём головного мозга новорождённой ящерицы может достигать 1/5 объёма тела, что отражает его значимость для молодого животного. 270 Поведение рептилий построено на врождённых реакциях, которые заметно индивидуализируются в процессе жизни. Этому способствует продолжительность приобретения индивидуального опыта. Крупные черепахи живут до 150-250 лет, змеи и ящерицы — 7-25 лет, а крокодилы — до 70-80 лет. У многих рептилий очень большую роль играет обучение. Они легко приручаются и быстро осваивают лабиринт или зрительные условные сигналы. Черепахи, принадлежащие к древнейшим анапсидам, могут вторично узнать фигуру, нарисованную сплошной линией, даже если она очерчена пунктиром или повёрнута на небольшой угол. Ящерицы и змеи способны к некоторому прогнозированию развития событий. Это позволяет им преследовать добычу уже после того, как она скрывается из виду. Однако водяные ужи, обученные правильно проходить Т-образный лабиринт, после линьки всё забывают. Аналогично ведёт себя шипохвост, который, привыкнув к новой территории, забывает её после кратковременного охлаждения. Крокодилы проявляют довольно изощрённую заботу о потомстве. Они готовят кучи из земли, листвы и веток, куда откладывают яйца. Пока зародыши развиваются, крокодилы косвенными способами контролируют температуру и охраняют кладку, а затем выкапывают вылупляющихся потомков. У рептилий известна иерархическая организация компактных групп. Доминирующей игуане уступают место не только на своей территории, но и на соседних, когда она направляется на охоту в общие нейтральные зоны. Иерархия у гигантских черепах Галапагосских островов выражается в строго установленном порядке следования друг за другом во время миграций. Групповая организация рептилий является одной из сложных форм общественного поведения, которая была достигнута у холоднокровных животных. Перечисленные особенности современных животных только косвенно отражают зоопсихологические достижения вымерших рептилий. Однако всё разнообразие их поведения базируется на головном мозге общего архетипа, который заслуживает отдельного рассмотрения. § 37. Ассоциативный центр мозга рептилий Рассмотрев общий план строения нервной системы, следует отдельно остановиться на новых принципах организации и работы мозга, впервые реализованных у рептилий. Нервная система архаичных амниот стала логическим развитием строения удачной амфибийной конструкции. Однако мозг амфибий практически выполнял функцию сложного рефлекторного аппарата, а его интеллектуальные возможности остались невостребованными. Эволюция амфибий решалась мышцами, зубами, линейными размерами и масштабами 271 Рис. III-8. Сагиттальный (а) и горизонтальный (б) гистологические срезы через голову ящерицы (Lacerta agilis)в конце эмбрионального развития. Относительный размер мозга в несколько раз больше, чем у амфибий. К моменту вылупления рептилии обладают эффективным вестибулярным аппаратом, зрением, слухом и обонятельной системой. 272 размножения. Шло элементарное освоение пищевых ресурсов, где для развития сложного поведения не было ни места, ни биологической необходимости. Со следами этого периода эволюции позвоночных мы сталкиваемся, пытаясь выработать условные рефлексы у различных представителей современных амфибий. Крайне низкая обучаемость и отсутствие долговременной памяти для накопления индивидуального опыта показывают, что сложных поведенческих задач перед древними амфибиями никогда в их эволюции не стояло. Особенности развития органов чувств и признаки сложного поведения рептилий базируются на особенностях структурной организации головного мозга. Мозг рептилий отличается от мозга амфибий как в количественном, так и в качественном отношении. До появления амниот стратегии поведения или реакция на конкретный раздражитель выбирались по принципу доминантности (см. рис. III-6, е). Этот принцип состоит в том, что выраженного крупного ассоциативного центра мозга у многих первичноводных позвоночных или амфибий нет (см. рис. III-6, e). Выбор формы поведения происходит на основании сравнения активностей примерно равноценных отделов мозга, обслуживающих различные органы чувств. Решающее значение играет уровень возбуждения мозговых аналитических центров одного из анализаторов. Представительство органа чувств, достигшее в мозге наибольшего возбуждения, и становится основной областью для принятия решения. После выбора одной из инстинктивных реакций происходит её поведенческая реализация. Этот процесс осуществляется под контролем того же простого сравнения доминантностей. Если в процессе осуществления реакции возникает новое раздражение, которое изменяет соотношение возбуждений органов чувств, то поведенческая реализация инстинктивного процесса останавливается. Каждая конкретная ситуация отличается от предыдущей, но задействуется тот же набор органов чувств. Если наибольшее возбуждение достигается в той же сенсорной системе, то поведение сохраняется, а если в другой, то изменяется. Поскольку абсолютно идентичные условия в естественной жизни практически не встречаются, поведение даже самых примитивных анамний будет бесконечно разнообразно. Следовательно, поведение каждой особи будет индивидуальным с довольно высокой динамической адаптивностью. Первые признаки ассоциативного центра появились ещё в мозге амфибий. У них таким центром мог стать средний или промежуточный мозг. Для этого были все основания. В промежуточном мозге находятся нейроэндокринные центры, контролирующие половое поведение, миграции и энергетический баланс организма анамний. Через 273 288 Возникновение мозга птиц Птицы представляют собой обособленный класс теплокровных, двуногих и обычно летающих амниот. Они покрыты специализированными роговыми придатками — перьями. Современные птицы лишены зубов, но обладают удлинёнными челюстями, которые образуют клюв. Наземное передвижение обеспечивают парные задние конечности. Передние конечности видоизменены в крылья, приспособленные для полёта. Большинство птиц способны летать. Они могут находиться в воздухе очень долго и перелетать на большие расстояния. Сезонные миграции могут составлять несколько тысяч километров. В стабильном климате многие птицы не мигрируют или перемещаются на небольшие расстояния. Летают не все птицы. Пингвины, киви и страусы не обладали этой способностью или утратили её вторично. Птицы — яйцекладущие животные. Все птицы откладывают яйца, но не все их насиживают и заботятся о птенцах. Для нормального развития зародышей необходима контролируемая температура окружающей среды. Это достигается насиживанием яиц, устройством защищённых гнёзд или созданием специальных конструкций, где температура поддерживается за счёт химических реакций распада. Поведение птиц крайне сложно и разнообразно, как и их внешний вид (рис. III-11). У птиц в разных пропорциях встречается инстинктивное и ассоциативное поведение и сохраняется способность к обучению на протяжении всей жизни. Многие птицы обладают хорошей памятью и могут осуществлять несложную инструментальную деятельность. Они легко обучаются решать простые задачи и подражать человеческому голосу. «Слова», произносимые птицами, являются эмоциональным символом ситуационного настроения, в котором они их запомнили, а не результатом рассудочной деятельности. В отличие от всех рассмотренных выше групп животных, птицы любят играть друг с другом или с различными предметами. Они первые среди животных смогли заниматься деятельностью, не приносящей непосредственной биологической выгоды. Мозг птиц обеспечивает возможность предвидения событий. Они рассчитывают своё поведение на несколько шагов вперёд, что уже характерно для высших млекопитающих и человека. Это сходство сложного поведения с поведением млекопитающих свойственно относительно небольшой группе попугаев, врановых, хищных и некоторых других видов. Поведение большинства птиц контролируется преимущественно врождёнными 289 Рис. III-11. Разнообразие птиц.  290 294 Столь подробное описание морфологических особенностей строения продиктовано уже упомянутой парадоксальной специализацией птиц. Скелет, покровы, кровеносная, выделительная и дыхательная системы имеют столько специфических особенностей, что их вполне хватило бы для гарантированного исчезновения любой другой группы позвоночных. Исключение составляет нервная система птиц. Во всех систематических группах птиц она построена удивительно однотипно. Анатомически различия обычно сводятся к размерам головного мозга (рис. III-12), а видовые особенности выявляются только на цитоархитектоническом уровне. Она столь же специализирована, как и другие органы, но обладает уникальным архетипом строения, который оказался пригодным для всех форм адаптивного поведения и сред обитания. |