Савельев. Происхождение мозга
 Скачать 1.82 Mb. Скачать 1.82 Mb.
|

|
§ 43. Нервная система и органы чувств птиц Нервная система птиц состоит из центрального и периферического отделов. Головной мозг птиц крупнее, чем у любых современных представителей рептилий. Он заполняет полость черепа и имеет округлую форму при небольшой длине (см. рис. III-12). Самый крупный отдел — передний мозг. Он состоит из двух полушарий с гладкой поверхностью или слабо обозначенными продольными углублениями. Эти углубления не являются истинными бороздами, а отражают границы слоёв внутренних ядер (рис. III-13, а, в).Только у попугаев можно заметить небольшое морфологическое обособление височной доли полушарий. Основной объём полушарий занимают подкорковые ядра, кора имеет рудиментарное строение и занимает небольшую часть верхней стенки мозга. Полушария переднего мозга простираются назад до контакта с мозжечком. Следствием разрастания переднего мозга назад, а мозжечка — вперёд является то, что промежуточного мозга снаружи совсем не видно, хотя его можно определить по выросту эпифиза. Эпифиз у птиц развит незначительно, а гипофиз достаточно крупный. Средний мозг сильно развит, но из-за бокового расположения имеет нехарактерную внешнюю морфологию. Передние выпячивания крыши среднего мозга сдвинуты латерально. Их часто называют зрительными долями (Lobi optici). Мозжечок состоит из массивной средней части (червя), пересекаемой обычно 9 извилинами, и двух небольших боковых долей, которые гомологичны клочку мозжечка млекопитающих. Задний и продолговатый мозг имеет два резких изгиба, обусловленных ориентацией и подвижностью головы птиц. Гистологическое строение головного мозга птиц существенно отличается от такового у других групп позвоночных. Уже у амфибий можно выявить старый и древний стриатум, септум и в зачаточном виде базальные ядра переднего мозга. У рептилий появляется новый 295 Рис. III-12. Головной и спинной мозг птиц. Головной мозг птиц обладает крайне стабильной анатомией. Он укорочен в рострокаудальном направлении, а на его поверхности видны только парные полушария переднего мозга, крупный мозжечок и передняя часть крыши среднего мозга. Внешняя форма головного мозга практически не отличается даже у отдалённых видов. а, в — хохлатый осоед (Pernis ptilorhynchus); б — серый гусь (Anser anser); г — павлин (Pavo cristatus). 296 стриатум, который становится доминирующей структурой переднего мозга (см. рис. III-12; III-13, а). Важно подчеркнуть, что новый стриатум возникает у птиц вопреки развитию зачаточного неокортекса рептилий. Иначе говоря, у рептилий получили значительное развитие структуры стриатума и септума переднего мозга. Они расположены в базальных частях переднего мозга птиц и развиты намного лучше, чем у рептилий (см. рис. III-13, а).Однако рептилии приобрели и зачаточные корковые структуры переднего мозга, которые сформировались в результате развития дополнительной (половой) обонятельной системы. Эти структуры стали выполнять у рептилий функции нового интегративного мозгового центра на базе переднего мозга. Казалось бы, дальнейшее развитие корковой системы переднего мозга гарантировало бы птицам необходимые поведенческие преимущества. Тем не менее этого не произошло. У птиц корковые структуры, впервые появившиеся в переднем мозге рептилий, носят откровенно рудиментарный характер. Эволюция корковых структур переднего мозга рептилий полностью остановилась у птиц. Зачатки палео-, архи- и неокортекса практически не играют роли в контроле поведения, поскольку обоняние у птиц развито намного меньше, чем у рептилий. У большинства современных птиц нет развитого обоняния, а вомероназальная система полового обоняния у них вообще отсутствует. По-видимому, в период отделения предков птиц из общего рептилийного ствола обоняние перестало играть для них какую-либо роль. Крупный передний мозг остался функционально невостребован и стал морфологическим субстратом для возникновения ассоциативных центров. Зачаточные корковые структуры не могли играть существенной роли на фоне «освободившихся» от своих функций огромных ядер стриатума и септума. Невостребованная нейронная система этих структур надолго обеспечила мозг птиц резервами памяти и возможностями развития сложного адаптивного поведения. Следы кортикальных структур переднего мозга птиц расположены только в дорсальной зоне полушария, а большую часть мозга занимают стриатум, септум и неостриатум. Функции ассоциативного центра в переднем мозге птиц выполняют чрезвычайно развитые структуры стриатума (рис. III-14, а).Однако историческое название «гиперстриатум» не отражает реального происхождения этого центра птиц. Ранее считалось, что гиперстриатум возник из стриатума и является его гомологом. Специальные исследования кинетики пролиферации и миграции нейробластов у птиц позволили установить, что гиперстриатум состоит из нейронов различного происхождения. 297 Рис. III-13. Цитоархитектоника головного мозга птиц на примере обыкновенной неясыти (Strix aluco). Буквы сечений на схеме мозга соответствуют обозначениям гистологических срезов. Микрофотографии гистологических срезов сделаны в соответствии с линиями, показанными на цветной схеме мозга с латеральной поверхности. Оранжевый цвет — передний мозг, красный — средний мозг, фиолетовый — мозжечок, синий — задний и продолговатый мозг. 298 Они по большей части мигрируют из латеральной (paleopallium)и новой коры (neopallium).Особенностью развития стриатума стало формирование структур ядерного типа, а не стратифицированных корковых образований. У птиц и рептилий разрастание стриатума приводит к экспансии дорсального внутрижелудочкового бугорка, который практически полностью занимает полость латеральных желудочков (см. рис. III-14, а).Гиперстриатум птиц (рептилии имеют его признаки) выполняет функции новой коры. Он представляет собой высший ассоциативный центр, который определяет принятие решений и является основной зоной хранения индивидуального опыта. Через стриатум осуществляются контроль за двигательными функциями и связь с лимбической системой (Northcutt, 1981). Спинной мозг птиц развит очень хорошо. Он образует большие утолщения в плечевой и поясничной области, откуда отходят нервы передних и задних конечностей (см. рис. III-12, а).В поясничном утолщении верхняя стенка спинного мозга расходится, и центральный канал расширяется в ромбовидный синус, покрытый сверху только мозговыми оболочками. Спинномозговые нервы соединяются у птиц корешками ещё в канале позвоночного столба и выходят между дугами или через отверстия слившихся позвонков. В плечевом сплетении обычно участвуют 4 нерва, реже 3, а у страусов — только 1 нерв. Для управления задними конечностями у птиц существует два сплетения: поясничное и седалищное, или крестцовое. Поясничное сплетение обычно состоит из 3, но у страусов может включать и 5 нервов. Седалищное сплетение у всех птиц состоит из 4 крестцовых нервов. Черепно-мозговые нервы у птиц представлены 12 парами, которые начинаются по отдельности. Обонятельный нерв (I) идет от нижней поверхности обонятельной доли, которая спереди и снизу примыкает к полушариям переднего мозга. Зрительные нервы (II) после хиазмы плавно переходят в средний мозг, а блуждающий нерв, как и у рептилий, идёт из черепа далеко назад, иннервируя сердце, лёгкие, пищевод и желудок. В отличие от рептилий, добавочный нерв (XI) представлен тонкой веточкой блуждающего нерва, а подъязычный (XII) нерв имеет корешки, отходящие как от продолговатого, так и от спинного мозга (Савельев, 2001). Симпатическая нервная система птиц построена по общему для всех позвоночных принципу. Однако у птиц в шейной части расположен большой симпатический нерв, часто называемый пограничным стволом. Он лежит в канале, образованном двукорневыми началами поперечных отростков позвонков. Участок этого нерва прилежит к сонным артериям, а далее кзади симпатический нерв находится в 299 Рис. III-14. Строение полушария переднего мозга и зрительных долей среднего мозга птиц на примере обыкновенной неясыти (Strixaluco, а)и сизого голубя (Columbalivia, б). Микрофотографии соответствуют линиям, показанным на цветной схеме мозга. Оранжевый цвет — передний мозг, красный — средний мозг, фиолетовый — мозжечок, синий — задний и продолговатый мозг. 300 канале, образуемом головками и бугорками рёбер, и только в поясничной области он лежит свободно. По строению органов чувств птицы имеют несколько отличий от остальных животных. Осязание многих птиц (кулики, утки) сосредоточено в челюстном аппарате, где расположены специализированные механорецепторы (тельца Гранди и Хербста). Осязательные тельца расположены в коже у основания крупных перьев и сконцентрированы в восковице, покрывающей основание клюва. Совы обладают особыми осязательными перьями, расположенными вокруг клюва. У дятлов, дроздов, попугаев и глухарей осязательные функции выполняет язык, весьма богатый нервными окончаниями; в нём присутствуют преимущественно различные типы механорецепторов, а не вкусовые сосочки. Птицы пользуются языком как органом осязания. Вкусовые рецепторы у большинства птиц развиты плохо, поскольку птицы обычно заглатывают пищу, не измельчая её в клюве. Зрение птиц чрезвычайно острое. Глаз способен к аккомодации и обладает рядом морфологических особенностей, позволяющих эффективно ориентироваться, успешно охотиться и издалека определять качество пищи. В сетчатке глаза птиц плотность фоторецепторов намного больше, чем у других позвоночных. В центральной части глаза расположен наклонный гребень, где сосредоточены светочувствительные клетки. Поскольку оптическая проекция растягивается на наклонной плоскости гребня, птицы видят эту часть изображения увеличенной в 0,25-1,2 раза. Дополнительным приспособлением, улучшающим цветовосприятие, являются масляные капли в колбочках сетчатки. Они функционируют, как цветовые фильтры, что позволяет различать больше оттенков в световом диапазоне электромагнитных волн. Зрительная система птиц позволяет воспринимать объекты, излучающие в ультрафиолетовом диапазоне, и поляризованный свет. Некоторые перелётные птицы могут непосредственно воспринимать направление электромагнитных полей, что позволяет им ориентироваться в любой точке на поверхности Земли. У птиц отлично развит слух. Орган слуха состоит из внутреннего, среднего и зачатков наружного уха. Птицы воспринимают звуковые сигналы очень широкого диапазона. В этом отношении они могут намного превосходить многих млекопитающих как по диапазону, так и по чувствительности к слабым звукам. У птиц хорошо развит вестибулярный аппарат. Он состоит из полукружных каналов, отвечающих за рецепцию, связанную с угловым ускорением, и гравитационного рецептора (рецептора линейного ускорения). Все эти компоненты есть и у рептилий. Однако у птиц появился совершенный мозговой центр анализа вестибулярных и кинестетических сигналов — мозжечок. 301 310 времени для адаптации крыльев к движениям в менее плотной среде. «Бег по воде» стал длительным биомеханическим этапом постепенного совершенствования специализированной мускулатуры и неврологических механизмов управления полётом птиц. Параллельно со становлением системы дыхания, кровообращения и мускулатуры передних конечностей у архаичных птиц возникла и теплокровность. Основной причиной её появления стала охотничья среда предков птиц. Значительное преимущество получали те животные, которые могли максимально долго пребывать в холодной воде (см. рис. III-15). Понятно, что теплокровность стала огромным достижением для птиц. Пребывая в холодной воде дольше, чем их конкуренты, они повышали вероятность добывания пищи и снижали общие энергетические затраты. С нырянием и плаванием в холодной воде, по-видимому, связан и тотальный переход предков птиц к откладке яиц. При этом способе размножения животные избавлялись от необходимости согревать и носить с собой своих будущих потомков. Это позволяло уменьшить массу тела животного, повысить плавучесть и снизить энергетические затраты при плавании и нырянии. Насиживаемые или охраняемые кладки яиц можно было увеличивать постепенно. Самки могли откладывать более крупные яйца, что привело к увеличению размеров молодых животных. Переход к плаванию и нырянию предопределил появление крыльев, развитых грудных мышц, исчезновение хвоста и развитие хвостовых желёз птиц. Перья, которые позволяли сохранять тепло при плавании и нырянии, нуждались в специальной защите от намокания. В противном случае архаичным птицам приходилось подолгу сушить своё оперение, как и некоторым современным птицам. Не исключено, что хвостовая железа появилась в эволюции птиц довольно поздно. Однако высока вероятность того, что это произошло одновременно с переходом к плаванию. Следовательно, птицы возникли в несколько этапов. На первом этапе это были небольшие оперённые рептилии, перешедшие к питанию рыбой на мелководье. В связи с новым типом питания протоптицы стали двуногими, а передние конечности долго оставались невостребованными. Обоняние было в значительной степени утрачено, а зрение стало доминирующим дистантным анализатором. На втором этапе переход к плаванию и нырянию привёл к развитию машущих передних конечностей и мощных грудных мышц. Охота в холодных водах и продолжительное ныряние стали стимулом для развития сложного дыхания, теплокровности и внеорганизменного развития эмбрионов. Передний мозг, утратив функции обонятельного анализатора, 311 стал структурной базой для развития памяти, обучения и ассоциативного мышления птиц. Сложные движения в трёхмерной среде стали причиной развития мозжечка птиц как сенсомоторного центра координации движений. На третьем этапе архаичные птицы перешли к «бегу по воде» при активной работе крыльев. Такой способ быстрого перемещения над водой стал предтечей свободного полёта и создал переходные условия для медленного становления неврологических систем контроля полёта. Свободный полёт возник после увеличения размеров крыльев и изменения геометрии маховых перьев. § 45. Адаптивная радиация птиц В довольно разнообразную группу современных птиц входят летающие и нелетающие птицы. Считают, что перья, пневматичный скелет, воздушные мешки, теплокровность и другие морфологические отличия птиц сформировались в связи с адаптацией к активному полёту, поэтому бескилевые нелетающие птицы и пингвины чаще всего рассматриваются как вторично специализированные группы. Предполагается, что первоначально они были летающими, а затем утратили способность к полёту после перехода к жизни в водной среде или на земле. По-видимому, ситуация с появлением пингвинов и нелетающих птиц намного более запутанная, чем представлялось ранее. Все современные птицы имеют нервную систему чрезвычайно сходного строения. Это говорит о том, что предками птиц была одна общая группа рептилий. Они были уже настолько неврологически специализированы, что дальнейшие преобразования мозга становились модификационными. Достигнутого уровня адаптивной специализации нервной системы архаичных птиц было достаточно для успешного освоения любой доступной среды обитания. Опираясь на строение нервной системы, можно предположить следующее развитие событий. Первоначальный архетип нервной системы птиц сформировался у нелетающих рептилий при охоте на мелководье (рис. III-16, а-в).Это событие произошло задолго до начала активного полёта. Крупный мозг, утративший обонятельные функции и специализированный по зрительно-ассоциативному типу, стал базовой структурой для появления вполне конкурентных, но не летающих животных. Вполне возможно, что часть таких архаичных птиц вернулась к жизни на поверхности земли. Они стали наземными насекомоядными или всеядными животными, которые никогда не были способны летать. Такие реверсии в эволюции птиц происходили неоднократно, но достоверно говорить о современных следах этих событий весьма затруднительно. Скорее всего единственным примером такого рода 312 315 Происхождение мозга млекопитающих Основные признаки млекопитающих (Vertebrata)— волосяной покров, молочные железы и теплокровность (рис. III-17). По меньшей мере два из этих признаков должны присутствовать у каждого представителя группы. У китов, моржей, броненосцев, голых землекопов и некоторых других млекопитающих волосяной покров развит незначительно или отсутствует. Основным отличием млекопитающих считают специальные железы, молоком которых самки выкармливают детёнышей. Млекопитающие имеют также сальные и потовые железы кожи. Эти железы ассоциированы с волосяным покровом и выполняют защитные, коммуникационные и репродуктивные функции. Молочные железы являются эволюционными производными специализированных кожных желёз. Только у млекопитающих есть зрелые эритроциты (красные клетки крови), лишённые ядра. У всех остальных анамний и амниот клетки крови имеют ядра. Столь же глубокие отличия млекопитающих связаны и с головным мозгом. В переднем мозге формируется шестислойная кора, а мозжечок состоит из червя и парных полушарий, которые имеют все млекопитающие (рис. III-18-III-20). Подобных образований головного мозга у других амниот нет (Савельев, 2001). § 46. Обзор классификации млекопитающих Класс современных млекопитающих разделяют на два подкласса: первозвери (Prototheria)и настоящие звери (Theria).К первозверям относят однопроходных млекопитающих (Monotremata), которые могут откладывать яйца, но выкармливают детёнышей молоком и имеют своеобразный волосяной покров. Настоящие звери объединяют две обширные группы низших зверей (Metatheria)и высших зверей (Eutheria).Низшие звери представляют собой сумчатых млекопитающих американского и австралийского происхождения, а к высшим зверям относят всех остальных млекопитающих, объединённых в 18 обширных и часто искусственных отрядов (см. рис III-17). Основным отличием низших зверей от высших является морфофункциональный тип организации внутриматочного питания эмбрионов и плодов. Низшие звери рождают очень небольших детёнышей, которые скорее напоминают эмбрионов. Их дальнейшее развитие проходит в специальной сумке, куда открываются молочные железы. Высшие млекопитающие рождают, как правило, крупных и более сформированных детёнышей. Развитие эмбрионов и плодов высших млекопитающих обеспечивается 316 Рис. III-18. Головной и спинной мозг современных млекопитающих. а — вскрытый головной мозг обезьяны с двумя утолщениями спинного мозга со спинной стороны; б — крот (Talpaeuropaea) с отпрепарированным мозгом; в — макака-резус (Масаса mulatta) с видом на мозг с латеральной поверхности; г — сагиттальный разрез головы свиньи (Sus scrofa). 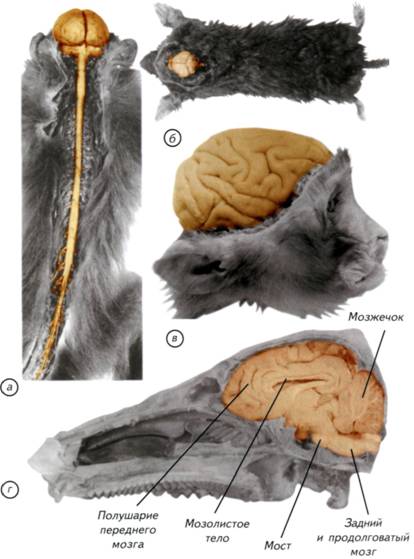 320 могут быть меньше 10 см и массой 50-90 г. Среди сумчатых большую группу составляют передвигающиеся на задних конечностях кенгуру (Macropodidae).У крупных кенгуру (Macropus, Megaleia)прыжки достигают 10 м, а скорость движения около 40 км/ч. Прыгая на задних конечностях, кенгуру используют хвост как балансир, что требует хорошей координации движений и контроля за центром тяжести тела. Этим задачам соответствуют развитый мозжечок и неокортекс. У большинства мелких сумчатых лиссэнцефальный (лишённый борозд и извилин) передний мозг. Хищные сумчатые (Dasyuridae), австралийские бандикуты (Peramelidae)и сумчатые медведи (Phascolarctidae)имеют развитую гирификацию или одиночные извилины. Насекомоядные (Insectivora)представляют собой наиболее примитивную в неврологическом отношении группу высших зверей. Строение их головного мозга сохранило множество архаических черт. Он лиссэнцефален, имеет небольшой неокортекс и развитые обонятельные центры (см. рис. III-18; III-19). Биология размножения насекомоядных традиционна и достаточно прогрессивна для млекопитающих. Она включает не только выкармливание детёнышей молоком, но и довольно продолжительную заботу о потомстве. По-видимому, насекомоядные специализировались на заре эволюции млекопитающих, что позволило им сохранить некоторые признаки наиболее древних черт морфологического строения. Надо подчеркнуть, что ежи (Erinaceidae), гимнуровые (Echinosoricidae), тенрековые (Tenrecidae)и кротовые (Talpidae)намного примитивнее в неврологическом отношении, чем современные сумчатые. Это делает их привлекательными объектами для эволюционной неврологии. Представители этой группы наиболее близки к архетипу строения мозга древнейших млекопитающих. Среди всего разнообразия млекопитающих, кроме насекомоядных, следует выделить грызунов и зайцеобразных, которые могут пролить свет на историю прогрессивного развития сложной архитектуры переднего мозга (см. рис. III-18; III-19; III-21). Вместе с тем ни в одной группе животных до млекопитающих мозг не достигал таких гигантских размеров, как у приматов, хоботных и китообразных (см. рис. III-19, г; III-22, б). Ни в одной группе позвоночных не было столь избыточного неврологического субстрата для развития памяти, индивидуального научения и ассоциативного мышления. В головном мозге высших млекопитающих сложились совершенно новые принципы иерархической обработки информации в распределённых системах, которые стали основой для мышления. Мозг высших млекопитающих отличается межполушарной системой связей — мозолистым телом. Это образование состоит из отростков нейронов, 321 Рис. III-19. Головной мозг современных млекопитающих. а — ёж (Erinaceus europaeus); б — кролик (Oryctolagus cuniculus); в — домашняя корова; г — афалина (Tursiops truncatus). Головной мозг кролика и ежа лиссэнцефальный, полушария переднего мозга коровы и дельфина содержат большое количество борозд и извилин. 322 323 Рис. III-20. Гистологические срезы через среднюю часть полушарий лиссэнцефального (а, б) и гирифицированного (б) переднего мозга. а — крыса (Rattus norvegicus); 6 — бобр (Castor fiber); в — медведь (Ursusarctos). 324 В связи с этим проблема происхождения млекопитающих сводится к поиску среды, которая удовлетворяла бы всем перечисленным требованиям. Она должна быть очень сложной, динамически изменчивой и весьма агрессивной. Для того чтобы представить себе эти условия возникновения млекопитающих, необходимо рассмотреть те неврологические приобретения нервной системы млекопитающих, которые отличают их от ранее рассмотренных амниот. |