Савельев. Происхождение мозга
 Скачать 1.82 Mb. Скачать 1.82 Mb.
|

|
§ 24. Эволюция ганглиозной нервной системы На заре эволюции многоклеточных сформировалась группа кишечнополостных с диффузной нервной системой (см. рис. II-4, а; рис. II-11, а). Возможный вариант возникновения такой организации описан в начале этой главы. В случае кишечнополостных отдельные нейроны объединены в элементарную нервную сеть, которая относительно равномерно распределена между внутренним и наружным слоем клеток. При всей простоте эта конструкция оказалась достаточно эффективной, и её обладатели дожили до настоящего времени. Обычная для Европы пресноводная гидра является классическим образцом поведения животного с такой нервной системой (см. рис. I-2). Из этой стартовой позиции начался гигантский эволюционный эксперимент по созданию эффективной системы управления организмом. 163 На первом этапе эволюции произошёл наиболее простой вариант усложнения нервной системы — концентрация нервных клеток в ганглии. Процесс объединения нервных клеток шёл различными путями: во-первых, вокруг глотки, что характерно для всех без исключения животных того периода, во-вторых, вдоль несимметричного тела. Рис. II-11. Эволюционные изменения расположения центров нервной системы от радиально- к билатерально-симметричным животным. На поперечных сечениях: нервные тяжи и комиссуры — голубые, ганглии и скопления тел клеток — синие. Стрелками вокруг сечений показаны направления слияния нервных стволов, одним и тем же цветом — последствия концентрации нервных элементов. а, б — кишечнополостные; в, г — плоские черви; д — членистоногие; e — позвоночные; ж — круглые черви. 164 По выделенным тяжам нервных клеток проще проводить общий для всего тела управляющий сигнал. Это произошло уже у кишечнополостных, и такие животные дошли до нашего времени. У многих современных актиний есть «скоростные» морфологически выделенные нервные стволы (см. рис. II-4, в).Появление таких стволов с признаками параллельных ганглиев произошло в результате объединения соседних нейронов. Для огромного большинства кишечнополостных такого усложнения диффузной нервной системы оказалось более чем достаточно. Они достигли того неврологического максимума, который при их образе жизни гарантировал питание и размножение. Однако даже среди радиально-симметричных кишечнополостных этот процесс происходил неодинаково. По-видимому, возникали как продольные, так и поперечные стволы из нервных клеток (см. рис. II-4, г, д). Судя по всему, достижением стало появление радиально-комиссуральной конструкции, которая могла сочетать в себе как дифференциальные сегментарные реакции, так и генерализованные ответы на раздражения (см. рис. II-5, в; II-11, б). Радиально-комиссуральная нервная система могла стать основой для появления любых сегментированных существ. Это связано с тем, что отдельные сегменты имели собственные комиссуры, что позволяло решать сегментарные «проблемы» на месте, за счёт автономных нервных механизмов. Организм из повторяющихся сегментов становится менее уязвимым. Способность к масштабной регенерации — не менее важное свойство, чем эффективность питания или размножения. Из такого организма могли произойти самые разнообразные билатерально-симметричные конструкции (см. рис. II-11, в, ж). Причиной этих процессов могли быть дальнейшая концентрация нервной системы в виде стволов и комиссур. В одном случае стволы могли сливаться дорсально, латерально и вентрально, как показано на рис. II-11, б зелёными стрелками. Результатом стало появление 4 продольных нервных стволов (см. рис. II-11, ж). Один ствол лежит дорсально, два — латерально и один — вентрально. Возникла своеобразная билатеральная симметрия, которая относительно редко встречается у других беспозвоночных. Такая нервная система характерна для нематод. По-видимому, комиссуральные тяжи у нематод исчезли уже вторично, в связи с упрощением их биологии (см. рис. II-5, б). Аналогичное, но альтернативное слияние нервных стволов показано фиолетовыми стрелками (см. рис. II-11). Оно могло произойти у ползающего в придонном слое организма. После слия- 165 Нервная система хордовых Палеонтологическая летопись содержит более чем скромные сведения о происхождении хордовых. Разрозненные и отрывочные данные не позволяют пока ясно представить себе события, которые привели к появлению первых хордовых, а затем и позвоночных животных. Все современные теории происхождения первых позвоночных построены на весьма сомнительных предположениях. В их основе лежат особенности строения современных условно «примитивных» видов, немногие ограниченные палеонтологические данные и столько же здравого смысла в сочетании с творческой фантазией. На таком сомнительном субстрате трудно создать достоверную гипотезу происхождения одной из ключевых групп животных, тем более что проверить её, по-видимому, никогда не удастся. В связи с этим имеют право на существование самые невероятные гипотезы эволюционного происхождения позвоночных. Любая из них может получить дополнительные доказательства по мере расширения наших палеонтологических знаний и технических возможностей. При создании настоящей теоретической реконструкции давно прошедших событий я старался опираться на основные принципы морфологических преобразований нервной системы. Воссоздание событий, приведших к появлению современных позвоночных, сводится к двум частным задачам. Первая состоит в том, чтобы обнаружить или представить себе древнее беспозвоночное животное, которое могло бы стать гипотетическим предком позвоночных. Вторая задача связана с реконструкцией событий, которые могли привести к появлению древних позвоночных. Для решения этих задач необходимо создать самый общий план строения позвоночных и беспозвоночных животных. Их сравнение позволит определить те морфологические проблемы, которые были решены в процессе эволюции. Современные позвоночные представляют собой весьма пёструю в морфологическом плане группу. У них билатеральная симметрия, хорда или позвоночник и дорсально расположенная центральная нервная система с полостью внутри (рис. II-12, б).Среди современных беспозвоночных также много билатерально-симметричных животных с дорсально расположенными головными ганглиями. Однако эти ганглии соединены комиссурами с нервной цепочкой, расположенной вентрально, под пищеварительной трубкой (см. рис. II-12, а).Морфология головных ганглиев беспозвоночных намного более разнообразна, чем строение головного мозга позвоночных животных 168 169 Рис. II-12. Общий план строения беспозвоночных и позвоночных. Схема нервной системы членистоногих (а) и позвоночных (б). Буквенное обозначение сечения (в) соответствует схеме в; г — увеличенный участок сечения ланцетника; д — сечение тела круглого червя. У круглых червей и ланцетника мышечные клетки формируют длинные отростки, которые доходят до нервных стволов или проникают в нервную трубку. Нейроны по этим отросткам передают управляющие сигналы к мышечным волокнам. 170 подход, который акцентирует внимание не на родственных связях животных, а на морфогенетических принципах эволюционных преобразований нервной ткани. С этих позиций можно провести оценку различных гипотез происхождения нервной системы хордовых. § 25. Теории происхождения хордовых Бытует несколько точек зрения на возникновение хордовых, которые различаются как по подходам к решению проблемы, так и по животным, избранным в качестве представителя предковых групп. Наиболее известны гипотезы возникновения хордовых от предковых форм членистоногих, паукообразных, немертин, кольчатых червей, иглокожих, гребневиков и кишечнодышащих. Для серьезного обсуждения пригодны только некоторые наиболее известные и обоснованные концепции. Тем не менее краткого упоминания заслуживают и весьма оригинальные идеи, если только они не обходят стороной проблему филогенетического становления нервной системы (Ливанов, 1955; Jollie, 1982; Иванов, 1991; Малахов, 1991). 172 кровеносная система внизу; направление тока крови у перевёрнутого червя и хордовых совпадает. Иначе говоря, чтобы из вентральной нервной цепочки и надглоточного ганглия, изображённого на рис. II-12, а, получить дорсальную нервную трубку с мозгом в одной плоскости (cм. рис. II-12, б), надо допустить множество невероятных событий. Для того чтобы обойти перечисленные затруднения, было предложено универсальное объяснение, которое предполагает дифференциальную редукцию наиболее «неудобных» для таких филогенетических построений отделов нервной системы. Этот подход возник ещё в конце XIX в. в связи со старой идеей о том, что предком позвоночных могло быть животное с вентральной нервной цепочкой и надглоточным ганглием. Это существо по загадочным причинам перевернулось к верху брюхом и стало ползать на спине. Понятно, что в этом случае надглоточный ганглий оказывается под пищеводом. Однако мозг у позвоночных находится не на груди. Таким образом, возникла проблема, связанная с «неудобным» положением головных ганглиев у перевернувшегося животного. Было сделано предположение, что головной специализированный ганглий беспозвоночных мог просто ненадолго редуцироваться, а затем возникнуть вновь. Его вторичное возникновение произошло в новом месте, уже соответствующем положению мозга хордовых. В несколько изменённом виде эта гипотеза продолжает разрабатываться в настоящее время (Berrill, 1955; Foreman etal., 1985; Jefferies, 1986; Малахов, 1991). Её сущность сводится к переносу гипотетического процесса переворачивания дорновского червя на эмбриологическую стадию. Считается, что в эмбриональном периоде мозг «небольшой», и можно легче допустить его редукцию. Это простодушное предположение привело к выведению вторичноротых из гипотетического диплеуруло-образного предка. Предполагается, что личинки кишечнодышащих — типичные диплеурулы, сходные по организации с предковыми формами хордовых. Согласно этой гипотезе рот и анус диплеурулообразного предка являются частями длинного щелевидного бластопора, а анус смещён на морфологически спинную сторону. У вторичноротых выделяют постнатальный (IV) отдел тела, снабжаемый целомической полостью от 3-й пары целомических карманов диплеурулы. На основании этого вывода телосома погонофор гомологизируется с IV отделом тела всех прочих вторичноротых. Исходя из этих рассуждений, предполагается, что «хордовые являются перевёрнутыми вторичноротыми», стебель Pelmatozoaгомологичен постнатальному отделу, а мадрепоровая плоскость иглокожих гомологична сагиттальной плоскости остальных вторичноротых. Гипотеза неотенического происхождения хордовых от 173 177 Рис. II-13. Внешний вид, общий план строения половозрелой асцидий и реконструкция головы свободноплавающей личинки. а, г — внешний вид и схема строения асцидий; 6 — ганглий половозрелой асцидий; в — схема строения головы и сечение личинки асцидий. У личинки нервная система представлена трубкой с асимметричным головным расширением и двумя нервными стволами. Один ствол лежит выше нервной трубки, а другой — ниже хорды. 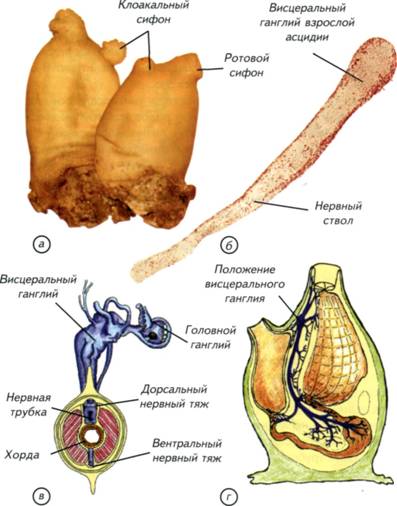 178 179 Рис. II-14. Внешний вид передней части нервной трубки и гистологические срезы через нервную систему и хорду ланцетника. а, б — две проекции реконструкции передней части нервной трубки ланцетника; в — микрофотографии клеток Рохда; г — светочувствительные и пигментные клетки; обзорная схема среза (д) и микрофотография участка тела ланцетника позади головной зоны (е); д — очерчен фрагмент, иллюстрированный микрофотографией на е.  180 183 Рис. II-15. Основные этапы (показано большими стрелками) возникновения хордовых. а, б — прикреплённое и свободноплавающее животное с радиальной симметрией; в-д — билатерально-симметричное животное с 4 нервными стволами; e — хордовое животное с продольно сливающимися ганглиями; ж, з — свободноплавающее хордовое с нервной трубкой и периферическими нервными ганглиями; д, ж — поперечные сечения тел животных; д — стрелками показано направление слияния нервных стволов.  184 185 Рис. II-16. Два варианта возможного возникновения нервной трубки (показано стрелками). В верхнем ряду показаны этапы формирования нервной трубки из 4 нервных стволов. Дорсальные и вентральные нервные стволы сливаются попарно, что приводит к формированию нервной трубки а, д, e, r. В нижнем ряду иллюстрировано появление нервной трубки из варианта организации с двумя нервными стволами а-г. Он предусматривает поворот животного на 90°, что менее вероятно, чем слияние дорсальных и вентральных нервных стволов. 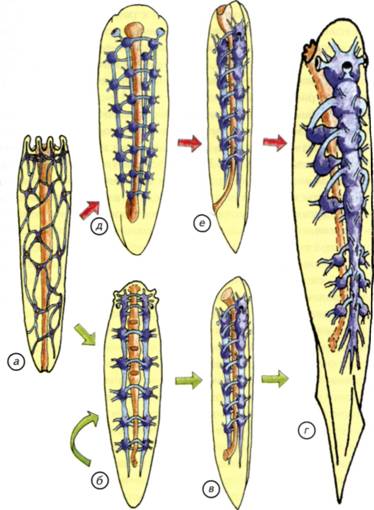 186 193 Первичноводные позвоночные Тип позвоночных (Vertebrata)включает в себя подтип бесчерепных (Acrania)и подтип черепных (Craniata).Единственными представителями бесчерепных являются многочисленные ланцетники (Amphioxi), которые рассматривались в предыдущем разделе, посвящённом происхождению позвоночных. Настоящий раздел охватывает обширную и разнообразную группу первичноводных черепных позвоночных. Они сформировались и долгое время морфологически изменялись в водной среде. В процессе эволюции именно этой группы сложилась нервная система, характерная для всех остальных позвоночных. Впервые возникло трубчатое строение нервной системы. Полости желудочков головного и спинного мозга стали непрерывными, появилась циркуляция внутрижелудочковой мозговой жидкости. В сочетании с эффективной кровеносной системой и внутренним скелетом это создало потенциальную возможность увеличивать размеры нервной системы первых водных позвоночных практически без ограничений. Эти эволюционные перспективы наращивания размеров мозга и параллельного увеличения сложности поведения не были реализованы в водной среде. Потребовались особые условия выхода на сушу, которые стали стимулом для развития новых качеств мозга и его ассоциативных функций (Northcutt, Davis, 1983). Однако благодаря первичноводным позвоночным нервная система приобрела те универсальные черты, которые послужили морфологической основой для дальнейшей эволюции. Его конструкция оказалась настолько эффективной, что по существу не подверглась принципиальным изменениям за всю историю эволюции позвоночных (Nieuwenhuys, 1982). Конечно, мозг адаптивно перестраивался во всех систематических группах, но принципы структурной организации центральной и периферической нервной системы сохранились без изменений. На конструкцию нервной системы радикально не повлияли появление ни челюстного аппарата, ни плавников, ни конечностей, ни выход на сушу, ни адаптация к полёту, ни теплокровность. Нервная система, как и позвоночник, оказалась тем органом, который обеспечивал выживание и прогресс всей группы позвоночных. Морфофункциональные отделы головного мозга, как и их основные функции, остаются неизменными на всём протяжении эволюции от миноги до человека. Эта универсальность строения нервной системы говорит об общности происхождения всех позвоночных. Делать предположения о полифилитичности происхождения позвоночных 194 198 Важнейшую часть центральной нервной системы первичноводных позвоночных представляет спинной мозг. Граница между спинным и головным мозгом достаточно условна, поскольку отростки клеток из головного мозга проникают в спинной и наоборот (Nieuwenhuys, 1998). Головной мозг, как правило, расположен дорсально и окружён хрящами или костями черепной коробки. Спинной мозг лежит в полости позвоночного канала, который образован невральными дугами позвонков (см. рис. II-19, а). У большинства позвоночных спинной мозг имеет однотипную гистологическую структуру. Тела нервных клеток (серое вещество) обычно расположены вокруг центрального канала в виде классической «бабочки», свойственной большинству амниот. У анамний картина несколько смазана, и расположение тел нейронов на разрезах спинного мозга в виде «бабочки» обычно не встречается (см. рис. II-19, а, б). Верхнюю часть серого вещества называют дорсальными (спинными) чувствующими рогами, а нижнюю — вентральными (брюшными) двигательными рогами спинного мозга. Спинные рога обычно содержат мелкие вставочные нейроны, а брюшные — крупные моторные. Через спинномозговые нервы осуществляются соматическая и висцеральная чувствительность, проходит соматически-двигательная и висцерально-двигательная иннервация. Соматическая чувствительность включает в себя рецепцию кожных, сухожильных, связочных и мышечных сигналов. Висцеральная чувствительность включает вкусовую рецепцию и сигналы от внутренних органов. Соматически-двигательная иннервация обслуживает скелетные мышцы, а висцерально-двигательная — железы, глоточную, лицевую мимическую и челюстную мускулатуру, гладкую мускулатуру кожи, сосудов и внутренних органов. В состав спинного мозга традиционно включают ганглий дорсального корешка спинного мозга, или спинальный ганглий, в котором расположены тела афферентных (чувствительных) соматических и висцеральных нейронов (см. рис. II-19, а, б). Эти клетки связаны со спинным мозгом коротким отростком, который оканчивается в дорсальных рогах серого вещества. Вентральная часть спинного мозга составлена из эфферентных (двигательных) волокон. Они начинаются от клеток, расположенных в вентральных рогах спинного мозга (Савельев, 2001). Не у всех позвоночных есть чёткое разделение корешков спинного мозга на моторные и сенсорные. У многих первичноводных позвоночных и амфибий висцеральные эфферентные волокна выходят из спинного мозга как через дорсальные, так и через вентральные корешки. У ланцетника, миног и миксин висцеральные эфферентные 199 Рис. II-19. Гистологические срезы через голову и мозг рыбы. 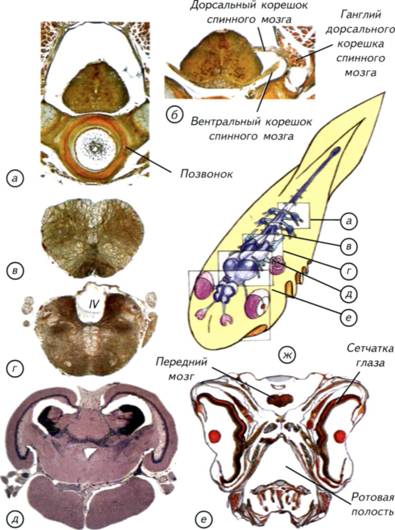 а — через спинной мозг и позвонок с невральными дужками; б — через спинной мозг в месте отхождения моторных и сенсорных нервов; в — через продолговатый мозг; г — через задний мозг; д — через средний мозг; e — через голову в зоне полушарий переднего мозга. Обозначения сечений на схеме ж соответствуют буквенным обозначениям срезов. IV— четвёртый желудочек. 200 201 Рис. II-20. Основные варианты строения мозга первичноводных. Прорисовки срезов через: F — полушария переднего мозга; G — промежуточный мозг; R — средний мозг; S — задний мозг. Уровень сечений соответствует обозначениям на схеме мозга с дорсальной поверхности (н).I—IV— номера мозговых желудочков. а — пластиножаберные и американский чешуйчатник; б — рогозуб; в — карпозубые; г — ильная рыба; д — американский чешуйчатник; e — амударьинский лопатонос; ж — колючая акула; з — миксина; и — ильная рыба; к — колючая акула; л — электрический скат; м — карась. Синим цветом отмечена наружная стенка мозга, красным — прижелудочковый слой клеток, зелёным — сосудистое сплетение. 202 203 Рис. II-21. Морфологические варианты развития головного мозга первичноводных позвоночных. Мозг сверху и боковая проекция головного и спинного мозга, вписанная в контур тела. а — у саргана преобладают зрительные центры среднего мозга; б — у рыбы-луны преобладают зрительные центры среднего мозга и нейрогормональный промежуточный мозг; в — у карпа доминирует представительство пищеварительной системы в заднем мозге; г — у акулы доминирует обонятельный передний мозг. 204 предопределяют способности пелагических акул к обнаружению пищи на больших расстояниях. При поступательном движении у большинства серых акул (Carcharhinidae)вода захватывается входной ноздрей, как воздухозаборником самолёта. Это обеспечивает быструю смену воды и возможность захвата пузырьков воздуха. Воздушные пузырьки могут захватываться и удерживаться в органах обоняния, если при движении со скоростью больше 1,2 м/с рострум приподнимается на 1-2 см над поверхностью воды. Пузырьки воздуха удерживаются в органах обоняния при помощи специализированных перегородок в обонятельном мешке — ламелл и снижения скорости движения животного. При задержке воздуха между ламеллами происходит растворение содержащихся в пузырьках веществ. Акулы не могут непосредственно рецептировать воздух и поэтому депонируют его в обонятельном мешке, дожидаясь повышения концентрации веществ вокруг пузырьков. Если вода не содержит привлекательных запахов, то акулы набирают скорость и выдавливают водой пузырьки воздуха, расположенные между ламелл, содержащих рецепторные клетки. Затем цикл повторяется. Надо отметить, что площадь поверхности рецепторных ламелл органов обоняния серых акул массой 75 кг может быть больше, чем у человека, в 60-90 раз. В воздухе над поверхностью океана запахи распространяются очень быстро, что позволяет акулам эффективно находить добычу. Следовательно, длиннокрылая акула может анализировать химические сигналы как из водной, так и из воздушной среды. Для обслуживания столь развитой обонятельной системы у серых акул сформировался гипертрофированный передний мозг, а обоняние стало ведущей системой афферентации. Если экстраполировать на человека образ мира такой акулы, то он предстанет как богатая запахами, но почти тёмная комната, она будет разделена по вертикали на мир водных и мир воздушных запахов. Через запахи будет передаваться почти вся информация, хотя кожные механорецепторы, специализированные электрорецепторные органы и весьма слабое зрение будут дополнять «обонятельную» картину мира (Halpern, 1980). Следует отметить, что различия в строении переднего мозга проявляются как на количественном, так и на качественном уровне (см. рис. II-20, F).Если у пластиножаберных и американского чешуйчатника (Lepidosiren paradoxe)полушария переднего мозга замкнуты (см. рис. II-20, а), то у рыбы-луны передний мозг заметно эвертирован (см. рис. II-20, в). Это означает, что полушария переднего мозга как бы вывернуты желудочками наружу. Некоторые костистые рыбы и рогозуб 205 (Neoceratodus forsterl)имеют промежуточный вариант строения полушарий, как показано на рис. II-20, б. В эвертированных полушариях переднее сосудистое сплетение лежит на верхней поверхности мозга или частично заполняет полость черепа. При обычном строении полушарий сосудистое сплетение располагается в полостях латеральных мозговых желудочков (Kappers, Huber, Grosby, 1936; Kardong, 1995). Промежуточный мозг не связан с конкретными анализаторами, за исключением пинеального комплекса. Последний включает в себя нейроэндокринные центры и теменной фоторецептор (глаз), который воспринимает только уровень освещённости и ответствен за суточные ритмы организма. В промежуточном мозге сосредоточены нейрогормональные центры, которые отвечают за рост, половое созревание, Рис. II-22. Крайние морфологические варианты развития мозжечка первичноводных позвоночных. Сечения мозга с боковой поверхности. а — минога, мозжечок морфологически не выделяется; б — форель, мозжечок увеличен, а его парус распространён под крышу среднего мозга в полость III желудочка; в — слонорыл из оз. Виктория (Африка). Его способность к электрорецепции вызывала гипертрофированное развитие аналитического центра — мозжечка, который полностью скрыл головной мозг. 206 осмотический баланс, работу эндокринных органов, сезонную и суточную активность животного. Через промежуточный мозг осуществляются связи между передним, средним, задним и продолговатым мозгом. У первичноводных животных уровень морфологического развития промежуточного мозга является своеобразным свидетельством определённой стратегии поведения животного. Если промежуточный мозг развит столь значительно, как у рыбы-луны, то механизмы регуляции поведения преимущественно гормональные (см. рис. II-21, б). Действительно, трудно найти среди первичноводных позвоночных животное с таким гипофизом, который лежал бы ростральнее переднего мозга, а обонятельные и зрительные нервы проходили бы над ним. У рыбы-луны гипертрофия промежуточного мозга сочетается с незначительными размерами переднего мозга и преобладанием зрительной системы. Эти животные ведут подвижную жизнь в мировом океане, им свойственна гигантская плодовитость. За один раз рыба-луна может отложить до 300 млн икринок. Рыба-луна питается зоопланктоном и практически не пользуется обонянием. Зрение ей необходимо в период размножения, когда животные собираются в большие скопления. Преобладание зрительных и гормональных центров над другими отделами мозга полностью соответствует биологии рыбы-луны. Таким образом, количественное доминирование нейроэндокринных центров промежуточного мозга над другими отделами центральной нервной системы является надёжным признаком гормонально-инстинктивного типа поведения животного (Foreman et al., 1985) . Зрительная система может стать ведущей системой афферентации у первичноводных позвоночных. В этом случае отмечается чрезвычайное развитие крыши среднего мозга — основной мозговой части зрительного анализатора. Примером такого строения мозга может быть средний мозг форели, чёрного марлина, летучей рыбы или плоскотелого саргана (см. рис. II-18, в, г, II-19, д; II-20, ж, и; II-21, а). Крыша среднего мозга образует крупные выпячивания, которые напоминают парные полушария переднего мозга, но это обманчивое впечатление исчезает, если рассмотреть III желудочек среднего мозга (см. рис. II-20, R). На сечениях видно, что реальных полушарий нет, а иллюзию создаёт симметричное латеральное расширение слоистой крыши (см. рис. II-19, д).Средний мозг у первичноводных животных является не только мозговым центром зрения. В крыше среднего мозга сосредоточены представительства боковой линии, вестибулярного аппарата и органов электрорецепции, которые хорошо развиты у многих первичноводных позвоночных. В вентральной части среднего мозга лежат преимущественно двигательные центры черепно-мозговых нервов. 207 215 Рис. II-23. Возможные этапы формирования основных отделов головного мозга позвоночных. а — наиболее примитивное состояние мозга с тремя сенсорными центрами; б — появление переднего мозга как центра для обработки внешних обонятельных сигналов; в — утрата связи аденогипофиза с глоточным эпителием, формирование парных полушарий переднего мозга и развитие этмоидной иннервации тройничным нервом. 216 собой и III желудочком только в каудальной зоне переднего мозга. Эта тенденция увеличения размеров обонятельных центров привела в конечном счете к формированию парных полушарий переднего мозга (см. рис. II-24, в). Однако наращивание количества нервных клеток в обонятельных центрах мозга первичноводных позвоночных шло различными путями. Ещё на заре возникновения переднего мозга были реализованы две различные эволюционные стратегии. Одна из них предполагала увеличение размеров переднего мозга путём размножения клеток в прижелудочковом слое с последующей их миграцией в наружные части стенки полушария. При этом типе развития мозга формируются парные полушария с замкнутой внешней стенкой. Такой вариант организации мы чаще всего встречаем у пластиножаберных (см. рис. II-24, в). У большинства костистых рыб и деление клеток, и их дифференцировка происходят в непосредственной близости от мозговых желудочков. Следствием такого типа дифференцировки становится выворачивание прижелудочковой поверхности полушарий наружу (см. рис. II-24, г).Эверсия полушарий переднего мозга стала тупиковым вариантом в эволюции нервной системы, если, конечно, считать выход на сушу эволюционно прогрессивным событием. У наземных позвоночных нет эвертированного варианта строения переднего мозга, а его обладатели не смогли покинуть водную среду. Продолжая реконструировать развитие основных отделов мозга, надо отметить, что с течением времени гипофизарный вкусовой карман утратил своё значение. Его функции стала выполнять часть ротовой полости, которая иннервировалась несколькими ветвями различных жаберных нервов (Mallatt, 1984). Такая множественность иннервации вкусовых клеток сохранилась и у современных позвоночных. Однако рецепторные и секреторные свойства клеток архаичной гипофизарной ямки не исчезли без следа. На её основе сформировался нейрогипофиз, который в сочетании с аденогипофизом стал ключевым органом регуляции желёз внутренней секреции. При переходе к активному плаванию нагрузка на сенсомоторные отделы возросла, что привело к увеличению размеров задних мозговых отделов, а потом и к образованию IV желудочка. Эти преобразования уже не были революционными, а возникший архетип мозга сохранился у всех современных позвоночных. Вполне понятно, что проведённая реконструкция возможных этапов эволюции первичноводных позвоночных носит гипотетический характер. Однако сравнительно-морфологические и палеонтологические данные косвенно подтверждают эту точку зрения (Stensio, 1963; Schaeffer, 1981). 217 Рис. II-24. Происхождение полушарий переднего мозга позвоночных. а — наиболее примитивное состояние переднего мозга с парными нервами, но одним полушарием; б — появление парных полушарий в результате роста центров, обслуживающих наружный хеморецептор; в — мозг современного животного и сечение через парные полушария; г — внешний вид и разрез через эвертированные полушария переднего мозга костистых рыб. Стрелками показаны пути эволюционных преобразований мозга. 218 § 29. Формирование мозга позвоночных Теперь попробуем предположить те события, которые могли привести к развитию описанной выше конструкции головного мозга древних первичноводных позвоночных. Жизнь в мелководной билатерали должна была иметь существенные преимущества для первых хордовых, в противном случае они бы никогда не появились. По-видимому, там небольшие и мягкотелые предки хордовых обнаружили неограниченное количество пищи, множество убежищ и идеальные условия для размножения. Надо уточнить, что перечисленные условия сохранялись достаточно долго, чтобы смогли сформироваться довольно сомнительные конкуренты беспозвоночных — предки хордовых. Вполне вероятно, что основная конкуренция в этих неглубоких водоёмах происходила между самими примитивными хордовыми без заметного вмешательства со стороны беспозвоночных. Первые хордовые были явными фильтраторами или собирателями придонных органических осадков. Наиболее эффективные преобразования ротовой полости могли ограничиваться созданием дополнительного ускорения протока воды через ротовую полость и жаберный аппарат. Этого можно было достигнуть как путём развития скелетно-мышечной системы жаберного аппарата, так и при помощи увеличения подвижности вентральной кожной складки (см. рис. II-23, в).Описанные события могли произойти ещё до развития скелетных конструкций в жаберной системе. Существование подобной дочелюстной системы захвата пищи или прокачивания воды могло стать причиной направленной дифференцировки тройничного, лицевого и подъязычного нервов. Основные эволюционные преобразования в этот период сводились к развитию систем, увеличивающих скорость сбора пищи и минимизирующих затраты на её поиск. Такие проблемы хорошо решаются в билатерали при помощи развития органов вкуса и обоняния. Именно эти события и предполагали формирование обоняния и его аналитического центра — переднего мозга. Параллельно формировался и орган вкуса. Первоначально это могла быть глоточная часть будущего гипофиза, а затем и специализированные ротовые и кожные вкусовые сосочки. С развитием дистантных обонятельных и вкусовых рецепторов возникла необходимость эффективно двигаться по направлению к источникам пищи. Для этого нужен мало-мальски эффективный механизм плавания. Сегментарная туловищная мускулатура беспозвоночных заложила основы для этих движений, но согласованность её сокращений сложилась вторично на базе сенсомоторных центров заднего и продолговатого мозга. 219 220 Рис. II-25. Основной план строения нервной системы и внешний вид возможного предка позвоночных. а — общий вид возможной нервной системы предка позвоночных; 6 — внешний вид гипотетического предка; в — реконструкция Arandaspis prionotolepis (по F. Knight); г — анаспид Birkenia из силура; д — цитаспид Anglaspis из раннего девона; e — птераспид Pterspis из раннего девона. Распространение бесчелюстных началось с билатерали, где они возникли. При освоении океана наиболее успешными оказались максимально защищенные формы, которые полностью исчезли. 221 222 Происхождение мозга наземных позвоночных Появление наземных позвоночных стало одним из самых занимательных сюжетов в эволюции. Конечно, трудно ожидать, что сравнительный анализ мозга современных амфибий и первичноводных позвоночных даст ответы на все интригующие вопросы этой обширной проблемы. Однако некоторые спорные аспекты происхождения наземных тетрапод можно рассмотреть с неврологических позиций. Этот подход может привести к формально непротиворечивым реконструкциям происхождения нервной системы первых наземных позвоночных. Основное внимание придётся уделить особенностям строения мозга современных амфибий, поскольку палеонтологические данные в отношении их нервной системы крайне скудны и ненадёжны. К современным амфибиям относят три относительно самостоятельные группы с довольно туманным прошлым; это хвостатые амфибии (саламандры и тритоны — Urodela), бесхвостые амфибии (лягушки и жабы — Anura)и безногие амфибии — червяги (червеобразные земноводные — Apoda)(рис. II-26). Хвостатые и бесхвостые амфибии внешне отличаются от рыб тем, что имеют рычажные конечности вместо парных плавников (см. рис. II-26). В личиночном состоянии почти все амфибии дышат жабрами, которые у ряда видов сохраняются в течение всей жизни. У взрослых животных обычно имеются лёгкие. Личинки амфибий развиваются в водной среде и обладают хрящевым скелетом, тогда как большинство взрослых животных обитает на земле и имеет скелет из костной ткани. Переход от личиночной стадии к взрослому животному часто связан с выходом на сушу, что приводит к значительным изменениям скелета и внутренних органов. У лягушек исчезает хвост, развиваются лёгкие вместо жабр и формируются конечности. Этот процесс носит название метаморфоза. Некоторые земноводные становятся половозрелыми на личиночной стадии. Это явление лучше всего известно для аксолотля и носит название неотении. У современных земноводных гладкая кожа, которая легко пересыхает. Амфибии размножаются преимущественно в воде и не имеют тазовых почек, поэтому нуждаются во влажной среде и в большинстве обитают поблизости от водоёмов. Несколько обособленное положение занимают червяги, которые обитают в почве или в водной среде. Они передвигаются в почве при помощи червеобразных движений и отличаются от других амфибий полным отсутствием конечностей и хвоста. Обитая в почве, они практически лишились зрения, хотя и обладают рудиментарными глазами (см. рис. II-26; рис. II-27-II-29). Среда их обитания наложила глубокий отпечаток на строение органов чувств и головной мозг. 223 Рис. II-26. Общий план строения нервной системы амфибий, вписанный в контур тела представителя каждого из отрядов. а — нервная система бесхвостых амфибий (Anura); б — нервная система хвостатых амфибий (Urodela); в — нервная система безногих амфибий (Apoda). Передний мозг и мозжечок более всего развиты у безногих амфибий в связи с почвенным образом жизни. Средний мозг наиболее совершенен у бесхвостых амфибий, а водные хвостатые амфибии сохранили признаки иннервации большинства органов боковой линии рыб. У многих безногих амфибий отсутствуют утолщения спинного мозга, маркирующие конечности, что свидетельствует об их отсутствии у предковых форм. 224 § 30. Особенности строения нервной системы амфибий 225 Рис. II-27. Головной и спинной мозг бесхвостой амфибии, основные нервы и микрофотографии гистологических срезов. Слева — общий вид головного и спинного мозга амфибий на примере лягушки; а-г — гистологические срезы, соответствующие сечениям, указанным на левом рисунке. На срезах римскими цифрами обозначены мозговые желудочки, а на схеме головного и спинного мозга — черепно-мозговые нервы. Спинномозговые нервы — п.spinI-XI. 226 235 Рис. II-30. Головной мозг амфибий и гистологические срезы через головной мозг и ретикулярные ядра среднего мозга. а — мозг Urodela; б — мозг Anura; в — мозг Apoda; г — горизонтальный срез через головной мозг шпорцевой лягушки (X. laevis) на уровне обонятельной и вомероназальной луковиц; д, e — ретикулярные ядра у лягушки (R. temporaria) и когтистого тритона (О. fisheri); г, e — импрегнация нитратом серебра, д — окраска по Маллори.  236 |