Савельев. Происхождение мозга
 Скачать 1.82 Mb. Скачать 1.82 Mb.
|

|
§ 47. Особенности нервной системы млекопитающих Центральная нервная система у млекопитающих развита больше, чем у какой-либо другой группы животных. Диаметр спинного мозга обычно несколько больше, чем у других тетрапод (см. рис. III-18, а). Он имеет два утолщения в грудном и поясничном отделах, которые связаны с поясами конечностей. В поясничном отделе люмбальный желудочек известен только у медведей. Мощные пучки восходящих волокон проходят по дорсальной поверхности спинного мозга млекопитающих. Признаки этих волокон впервые появляются у рептилий и птиц, но максимальное развитие они получают только у млекопитающих. Задний мозг млекопитающих хорошо развит и имеет крупный мозжечок. В отличие от мозжечка рептилий и птиц мозжечок млекопитающих состоит из центральной части, или червя, и двух боковых полушарий. Наиболее древней частью мозжечка является червь. Складчатость червя мозжечка хорошо развита у птиц. У млекопитающих по бокам червя мозжечка формируются парные полушария. Однако на развёртках поверхности мозжечок выглядит, как непрерывная и однородная структура. Тем не менее сохранилась старая морфологическая традиция выделять центральную (осевую) часть, называемую червём, и парные полушария мозжечка. На самом деле это только анатомические складки единой поверхности мозжечка. Площадь поверхности мозжечка может достигать огромных размеров. У человекообразных обезьян и многих копытных размеры поверхности борозд и извилин мозжечка могут быть больше, чем коры переднего мозга. Между полушариями мозжечка образуется система связей, которая позволяет клеткам коры мозжечка обмениваться сигналами, приходящими как с правой, так и с левой стороны тела. Эти волокна выходят на наружную поверхность мозга млекопитающих и меняют его анатомическое строение. В результате на базальной поверхности заднего мозга млекопитающих появляется мост, который отсутствует у других амниот (см. рис. III-18; III-19). У сумчатых, грызунов и насекомоядных размер червя не уступает каждому из полушарий, тогда как у приматов и хищных полушария мозжечка больше. 325 Рис. III-21. Внешний вид и разрезы головного мозга однопроходных, трубкозубов, беличьих и приматов (трубкозуб по Edinger, 1911). а, г — проехидна (Zaglossus); б — трубкозуб (Orycteropus afer); в — белка (Sciurus vulgaris); д — макака-резус (Macaca mulatta). У однопроходных может быть гирифицированный неокортекс, но отсутствует мозолистое тело, соединяющее межполушарными волокнами неокортекс переднего мозга. У плацентарных могут отсутствовать извилины, но мозолистое тело развито очень хорошо. 326 Надо подчеркнуть, что мозжечок не является ни сенсорным, ни моторным образованием. Он стоит в стороне от главных входных и выходных путей и координирует двигательную активность с нервной деятельностью других отделов головного мозга. Одной из важнейших функций мозжечка является интеграция различной соматической информации, поступающей от поверхности тела млекопитающего. Мозжечок также выполняет функции сенсомоторной координации, регуляции мышечного тонуса и поддержания равновесия, входит в состав двигательной системы, но не имеет прямых связей с чувствительными клетками и мускулатурой. Это своеобразная «надстройка» над восходящими и нисходящими путями и центрами ромбовидного мозга. Мозжечок низших позвоночных соединён с задним мозгом непрерывной пластинкой, в которой пучки волокон анатомически не выделяются. У млекопитающих эти пучки формируют три пары столбообразных структур — ножки мозжечка. Через ножки осуществляются связи мозжечка с остальной центральной нервной системой. Задние ножки состоят преимущественно из афферентных волокон, которые приходят из спинного мозга. В средние ножки входят волокна, соединяющие мозжечок и кору переднего мозга. Передние ножки сформированы из эфферентных волокон, соединяющих мозжечок и средний мозг. Огромный мозжечок млекопитающих возник из-за крайней необходимости анализа сенсомоторной информации и координации движений. Для обработки сенсомоторных и соматических сигналов сформировалась сложная кора мозжечка, которая сопоставима по размерам с полушариями переднего мозга. Средний мозг представляет собой плотное небольшое тело в виде двух пар возвышений, которые традиционно носят название четверохолмия (рис. III-22, г). У рептилий и амфибий крыша среднего мозга состоит из двух возвышений, в основном связанных с сетчаткой глаза. У млекопитающих они сохраняют свои функции и расположены в передней зоне четверохолмия. Появление задних бугорков среднего мозга обусловлено развитием слухового аппарата млекопитающих. Они являются слуховыми центрами и отражают уровень развития слуховой системы у конкретного вида. По соотношению бугорков четверохолмия можно судить о преобладании одного из органов чувств. У копытных и хищных наиболее развиты передние бугорки четверохолмия, которые связаны со зрением, а у китообразных, рукокрылых и утконоса преобладают слуховые задние бугорки. Однако крыша среднего мозга у зверей играет второстепенную роль в обработке как зрительной, так и слуховой информации. Роль ведущих центров начинают выполнять специализированные поля новой коры. Промежуточный мозг полностью 327 Рис. III-22. Гистологические срезы через мозг тупайи, дельфина, неокортекс бородавочника и ствол мозга кролика. а — тупайа (Tupaiatana); б — афалина (Tursiopstruncatus); в — неокортекс бородавочника (Phacochoerus aethiopicus); г — мозжечок, четверохолмие крыши среднего мозга и таламус промежуточного мозга кролика (Oryctolaguscuniculus). ВБ — верхний бугорок; НБ — нижний бугорок. 328 скрыт под полушариями переднего мозга. В нём хорошо развит эпифиз, а воронка, как и у всех позвоночных, завершается крупным гипофизом. Этот отдел выполняет преимущественно нейроэндокринные функции и регулирует половую активность млекопитающих. Внутри головного мозга находится сложная система мозговых желудочков. Полость спинного мозга переходит в ромбовидную ямку продолговатого мозга, которая носит название IV мозгового желудочка. Его передний край у большинства млекопитающих прикрыт мозжечком. Кпереди от мозжечка IV желудочек переходит в сильвиев водопровод — канал, проходящий под средним мозгом и ведущий в III желудочек. Ill желудочек представляет собой довольно узкое пространство между внутренними стенками промежуточного мозга. Спереди он сообщается с двумя боковыми желудочками через монроевы отверстия. I и II желудочки лежат внутри полушарий переднего мозга. Впереди III мозгового желудочка вследствие местного срастания полушарий вокруг треугольного пространства образуется так называемый V мозговой желудочек. Этот желудочек свойствен только млекопитающим. Разветвлённая система желудочков даёт возможность эффективно поддерживать водно-солевой обмен головного мозга (см. § 5, 7). Основным отличием строения мозга млекопитающих от мозга рептилий и птиц являются крупные парные полушария переднего мозга и мозжечка (см. рис. III-18; III-19). Поверхность полушарий переднего мозга млекопитающих может быть гладкой — лиссэнцефальной или складчатой — с бороздами и извилинами. Полушария переднего мозга млекопитающих разрастаются вбок и кзади, покрывая собой промежуточный мозг, часто также средний мозг, а у приматов и китообразных — продолговатый мозг (см. рис. III-19; III-22). Полушария переднего мозга имеют неправильную овальную форму и более или менее расширены сзади. У многих млекопитающих самых разнообразных групп (насекомоядные, грызуны, зайцеобразные) полушария гладкие, без борозд и извилин. У большинства высших млекопитающих в коре формируются борозды и извилины (см. рис. III-19-III-21). Однако извилины бывают развиты в различной степени даже в пределах одного отряда. Мозг низших приматов практически лишён борозд и извилин, тогда как у высших приматов они сходны с извилинами мозга человека (см. рис. III-22). С увеличением размеров полушарий непосредственно связано появление мозолистого тела — обширной зоны волокон, соединяющих левое и правое полушария. Это образование характерно для мозга высших млекопитающих, но отсутствует в мозге однопроходных и сумчатых (см. рис. III-18-III-21). 329 330 организацию. Кора переднего мозга млекопитающих имеет сходное строение со слоистой крышей среднего мозга низших позвоночных. Неокортекс млекопитающих имеет шестислойное строение. У взрослых млекопитающих различных видов кора может значительно различаться как по толщине, так и по выраженности отдельных слоёв (см. рис. III-22). Известно, что у животных с лиссэнцефальным мозгом поперечник коры больше, чем у животных с полушариями переднего мозга, имеющими борозды и извилины. Так, у бобров поперечник коры несколько больше, чем у человека и медведя, а у дельфина афалины меньше почти в 2 раза. При всём разнообразии вариантов строения неокортекса шестислойность коры млекопитающих не вызывает сомнения (см. рис. III-22; рис. III-23-III-26). I — молекулярный слой (lamina molecularis, lamina zonalis) — самый наружный слой коры, который содержит редкие клеточные тела. II — наружный гранулярный слой (lamina granulans externa) состоит из небольших и плотно расположенных клеточных тел. В моторных областях этот слой может включать небольшие пирамидные клетки, которые распространяют свои волокна на поверхности мозга или в молекулярном слое. III — наружный пирамидальный слой, или слой наружных пирамид (lamina pyramidalis externa). В этом слое преобладают пирамидные нейроны, которые крупнее во внутренних зонах слоя и мельче — в наружных. IV — внутренний гранулярный слой (lamina granulans interna). Этот слой состоит из плотно расположенных как пирамидных, так и непирамидных клеток. V — пирамидальный слой (lamina pyramidalis). В этом слое расположены преимущественно клетки округлой формы, которые посылают свои волокна к пирамидным нейронам. VI — слой полиморфных клеток (lamina multiformis). В нём находятся веретеновидные, округлые и модифицированные пирамидные клетки. В неокортексе млекопитающих есть пирамидные клетки, которые отсутствуют в мозге других позвоночных. В моторной коре находятся особенно крупные пирамидные нейроны, носящие название клеток Беца (см. рис. III-24). Эти нейроны посылают свои длинные отростки к мотонейронам спинного мозга и в двигательные ядра черепно-мозговых нервов. Их гигантский размер предопределён масштабами метаболической и функциональной активности. 331 Рис. III-23. Основные эволюционные приобретения мозга млекопитающих на примере домашней собаки (Camsfamiliaris). Неокортекс с развитыми извилинами и появление распределённых морфофункциональных полей на поверхности полушарий сняли ограничение на увеличение размеров мозга (а). Формирование развитого мозжечка с огромными парными полушариями решило проблему дифференциальной соматической чувствительности и координации положения тела в пространстве (б). 332 333 Рис. III-24. Головной мозг кролика (Oryctolagus cuniculus)снанесёнными на поверхность полушария основными функциональными полями неокортекса (а) и цитоархитектоника моторных и сенсорных полей коры. Гистологический срез через моторное (б) и зрительное поле (в) неокортекса. 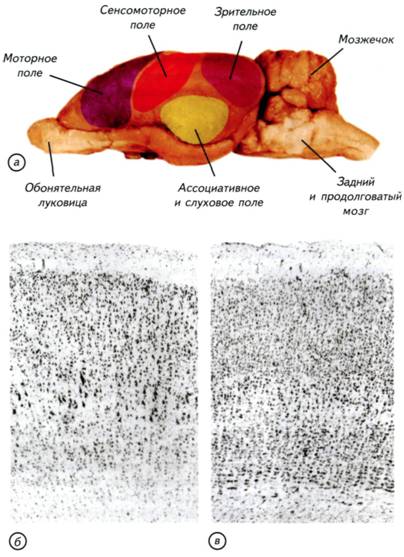 334 335 Рис. III-25. Дорсальная поверхность головного мозга млекопитающих. а — кролик (Oryctolagus cuniculus); б — шимпанзе (Pantroglodytes); в — лев (Pantheralео); г — обыкновенная игрунка (Callithrixjacchus). Уровень развития поведения млекопитающих только отчасти коррелирует с организацией борозд и извилин в полушариях переднего мозга. Значительную роль в гирификации играют размеры кортикальных полей различных сенсорных систем. Это маскирует реальные ассоциативные возможности вида. Мозг кролика и примата может быть лиссэнцефален, тогда как специализированный хищник (лев) может обладать гирифицированным мозгом, как у «интеллектуального» примата (шимпанзе). 336 его частью. Функциональные зоны коры имеют упорядоченное внутреннее строение. В каждой зоне или поле можно выделить своеобразные модули вертикальной упорядоченности организации коры. В самом обычном случае модуль представляет собой колонку или клубочек, в который включаются клетки, расположенные по всей толщине коры. Колонкой минимального размера обычно считают группу из 110 нейронов, расположенных между парой капилляров, проходящих через поперечник коры. Клетки колонок связаны с обработкой конкретного частного сигнала и рассматриваются в качестве локального функционального модуля. Более низкий уровень организации уже связан с межклеточным взаимодействием на уровне нейронов. Для неокортекса известны закономерности появления шипиков на отростках нейронов при развитии животных в обогащённой среде или при обучении. Чем богаче впечатлениями среда, в которой развивается млекопитающее, тем больше межклеточных контактов возникает между нейронами коры. Существование субстрата, связанного с сохранением индивидуального опыта, показывает, что возникновение неокортекса было ключом к расширению приспособительных возможностей млекопитающих. § 48. Органы чувств млекопитающих Органы чувств млекопитающих имеют ряд особенностей, которых нет у позвоночных других групп. У млекопитающих хорошо развит орган обоняния. В отличие от птиц он состоит из основного органа обоняния и вомероназальной системы. Основной орган обоняния используется для традиционных типов хеморецепции, а вомероназальная система (якобсонов орган) — для рецепции половых запахов. Развитие системы полового обоняния рептилий стало причиной возникновения корковых образований переднего мозга (см. § 39). Эти небольшие скопления клеток у поверхности полушарий переднего мозга не предвещали своим обладателям особых неврологических перспектив. Однако именно у млекопитающих эти структуры стали морфологической основой для возникновения шестислойной коры. Развитие коры переднего мозга свидетельствует о том, что у архаичных млекопитающих обоняние, особенно на первых этапах, играло основную роль среди дистантных хеморецепторов. Слуховая система млекопитающих хорошо развита благодаря появлению наружной слуховой раковины, усложнению строения улитки, развитию среднемозгового и коркового представительства акустического анализатора. У млекопитающих улитка впервые закручивается в спираль, которая у китообразных делает 1,5 оборота, у человека — 337 Рис. III-26. Цитоархитектоника слуховых полей неокортекса. 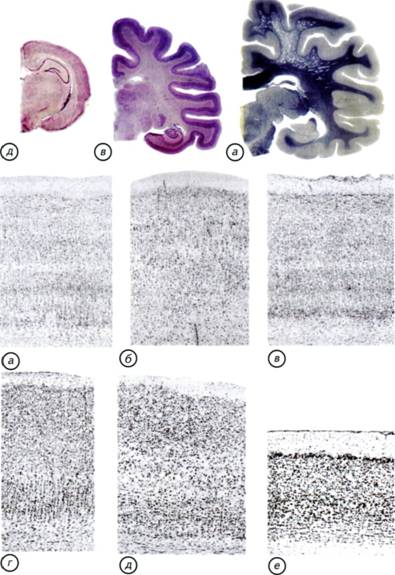 а — орангутан (Pongo pygmaeus); б — шимпанзе (Pantroglodytes); в — гиббон (Hylobateslar); г — кошка (Felis catus); д — крыса (Rattus norvegicus); e — крот (Talpa europaea). Обозначения срезов через левое полушарие соответствуют обозначениям участков коры. 338 343 Рис. III-27. Происхождение млекопитающих.  Рептилийные предки млекопитающих, покинув карбоновые растительные завалы, перешли к обитанию на ветвях деревьев (а-в). Переход к обитанию на деревьях стал ключевым событием для начала эволюции мозга млекопитающих. Ветви деревьев стали переходной средой для развития сенсорных и моторных систем млекопитающих. В кроне деревьев у архаичных млекопитающих сформировалась соматическая рецепция, построенная на свободных нервных окончаниях, оплетающих волосяные сумки и стержни волос (г). Развитие соматической чувствительности, пространственного зрения и системы сенсомоторной координации движений привело к формированию коры переднего мозга и парных полушарий мозжечка. В коре переднего мозга сформировалось представительство соматических, сенсомоторных, зрительных, вкусовых и слуховых центров. Эти неврологические приобретения позволили архаичным млекопитающим освоить сушу и водную среду (в, д, ж-н, р). На всех этапах эволюции млекопитающих происходили «возвраты» к наземным формам жизни. Одними из первых «спустились с деревьев» однопроходные, а затем — сумчатые (в, д, е, л). Значительно позднее к ним присоединились плацентарные млекопитающие. Рукокрылые с деревьев перешли к полёту, а приматы в основном сохранили свою связь с деревьями. 344 почвенных млекопитающих (голые землекопы) редуцируется даже волосяной покров. Маловероятно, что развитая соматосенсорная система и волосяной покров могли возникнуть у животных, ведущих полуподземный образ жизни. По-видимому, рептилийные предки млекопитающих, покинув карбоновые завалы, переместились в кроны деревьев (см. рис. III-27, в, г).Вертикальная «миграция» из плохо освещённых растительных буреломов в сумеречный мир крон деревьев выглядит вполне естественной. Этот переход не был радикальным изменением биологии рептилийных предков млекопитающих. Сохранились аналогичная трёхмерная жизненная среда и значение уже хорошо развитого вестибулярного аппарата. Вполне вероятно, что переход из нижних уровней карбоновых лесных завалов в крону деревьев происходил неоднократно, но с разными результатами. Только после появления первичной специализации головного мозга рептилий по обонятельному типу смогли сложиться необходимые предпосылки для формирования «древесной» группы архаичных млекопитающих. В сумеречных кронах деревьев необходим именно тот набор нейросенсорных, аналитических и репродуктивных приобретений, который известен у современных млекопитающих. Жизнь макросматиков в кронах деревьев практически исключала размножение в гнёздах или дуплах. Для небольших животных с развитым обонянием чужая кладка яиц была и остаётся идеальной и доступной пищей, поэтому доставшееся от рептилийных предков живорождение получило дальнейшее развитие. Было необходимо максимально продлить внутриутробное развитие эмбриона. Это позволяло избежать формирования гнезда и привязки к конкретной территории. Мать перемещалась за пищей вместе с детёнышем, что повышало вероятность их выживания. Наиболее простой способ увеличения продолжительности внутриутробного развития связан с отказом от питания эмбриона за счёт желтка. Запасы желтка нельзя бесконечно увеличивать в материнской матке. Намного эффективнее использовать простой диффузионный обмен кислорода, воды и метаболитов между стенкой желточного мешка и маткой. По-видимому, этим способом и была решена проблема внутриутробного развития архаичных млекопитающих. Древесные предки млекопитающих были весьма некрупными животными. Это позволяло им при помощи желтковой плаценты доращивать эмбрионы до вполне жизнеспособных размеров. Похожую репродуктивную стратегию используют современные сумчатые. Однако их желтковая плацента позволяет вырастить только небольшой эмбрион, который надо переводить в сумку с молочными железами. Поскольку архаичные млеко- |