Рецензенты кандидат химических наук В. Г. Коробко доктор биологических наук В. А. Гвоздев Патрушев Л. И
 Скачать 5.83 Mb. Скачать 5.83 Mb.
|

48 ч после трансфекции. Далее внутриклеточный уровень плазмидной ДНК начинает постепенно снижаться из-за ее цитопатического действия и гибели клеток. В результате системы, основанные на клетках COS, не могут быть использованы для крупномасштабной наработки рекомбинантных белков в течение длительного промежутка времени. Максимальное внутриклеточное содержание рекомбинантных белков в этих системах наблюдают через 72 ч после трансфекции, и их синтез продолжается в течение последующих 5–10 дней на фоне медленного снижения количества клеток в культуре, что позволяет все же использовать клетки COS для препаративного синтеза рекомбинантных белков. С этой целью трансфекции одновременно подвергают до 108 клеток с последующим их выращиванием на роллерах или микроносителях, сопровождаемым многократным сбором культуральной жидкости. Такой подход позволяет получать до нескольких миллиграммов рекомбинантного белка из вышеописанного пула трансфецированных клеток.7.4.Эукариотические системы экспрессии рекомбинантных генов, основанные на культурах клетокВыделение любого нового рекомбинантного гена описанными выше методами неизбежно заканчивается попытками получения его полноценной экспрессии в искусственных генетических системах. Только на первый взгляд экспрессия рекомбинантных генов в бактериальных клетках под контролем хорошо изученных регуляторных элементов бактерий представляется наиболее простой задачей. На практике, как уже обсуждалось ранее, экспрессия эукариотических генов в бактериях происходит крайне неэффективно из-за образования нерастворимых телец включения и отсутствия необходимых посттрансляционных модификаций рекомбинантных полипептидных цепей. Для преодоления этих затруднений в последнее время широко используются культивируемые клетки животных и растений, в геном которых рекомбинантные гены вводятся с помощью трансфекции. Однако эффективный синтез рекомбинантных белков зависит не только от используемых клеточных линий. Основное влияние на этот процесс оказывают конструкция экспрессирующего вектора, а также сам метод введения рекомбинантных ДНК в эукариотические клетки. В последнем случае низкая эффективность доставки рекомбинантных молекул в клетки и неоптимальная локализация сайтов их интеграции в геном могут свести на нет все достоинства клеточных линий и экспрессирующих векторов. Для повышения уровня экспрессии рекомбинантных генов, стабильно интегрированных в геном культивируемых клеток, разработаны методы их селективной амплификации. С использованием одного из таких подходов получены мутантные сублинии клеток яичников китайских хомячков CHO, в которых амплификация происходит в присутствии селектирующего агента, например метотрексата. Другие системы амплификации основаны на подавлении экспрессии жизненно важных ферментов, например глутаминсинтетазы или аденозиндезаминазы, под действием специфических ингибиторов: метионинсульфоксимина или 2’-дезоксикоформицина соответственно. Механизмы амплификации генов под действием ингибиторов ферментов будут подробнее рассмотрены ниже на примере метотрексата (см. раздел 7.4.1). Клеточные линии и клоны, стабильно продуцирующие рекомбинантные белки, как правило, получают путем отбора из пула гетерогенных клеток. И, наконец, еще одним существенным условием успешной экспрессии рекомбинантных генов является оптимизация самого процесса культивирования клеток, включая тщательный подбор культуральных сред. Для получения рекомбинантных белков кроме вышеупомянутого метода стабильно трансфецированных линий клеток используют еще два современных подхода. С целью наработки небольшого количества рекомбинантных белков и контроля функциональной целостности генно-инженерных конструкций часто применяют временно экспрессирующиеся векторы, которые обладают способностью к репликации в культивируемых клетках COS во внехромосомном состоянии. В этом случае источником рекомбинантных белков являются супернатанты клеток. При другом подходе рекомбинантные белки синтезируют в культивируемых клетках насекомых после их заражения векторами на основе генома бакуловирусов. Последний подход становится все более популярным, так как допускает стабильную экспрессию многих рекомбинантных генов, продукты которых могут находиться как внутри клеток, так и в ассоциированном с поверхностью клеток состоянии, или они могут секретироваться в культуральную жидкость. Недавно швейцарскими исследователями (С. Гайссе с соавт., 1996 г.) было проведено сравнительное изучение эффективности экспрессии гена цитокина человека – фактора, ингибирующего лейкозы (leukemia inhibitory factor – huLIF) в разных эукариотических системах. Фактор huLIF представляет собой белок среднего размера, полипептидная цепь которого содержит несколько потенциальных сайтов N-гликозилирования и в процессе фолдинга образует три дисульфидные связи. Поскольку полученные результаты представляют большой практический интерес для генной инженерии и биотехнологии, так как иллюстрируют многие основные принципы функционирования рекомбинантных генов в гетерологичных системах экспрессии, они будут кратко рассмотрены ниже. 7.4.1.Клетки яичников китайских хомячков (линия CHO)Эта линия клеток и ее многочисленные производные часто используются для синтеза рекомбинантных белков после предварительной эндогенной амплификации соответствующих рекомбинантных генов, введенных в клетки с помощью трансфекции. Спонтанная амплификация генов в нормальных клетках животных является редким событием, а в трансформированных (опухолевых) клетках она происходит с частотой 10-4–10-6 за клеточную генерацию. Частота амплификации определенных генов может быть повышена в результате некоторых экзогенных воздействий, включая инкубацию клеток с гидроксимочевиной, афидиколином, канцерогенами, а также под действием ультрафиолетового света или -излучения и в условиях гипоксии. Амплификация гена обычно начинается с его вырезания из хромосомы в составе протяженного сегмента ДНК, сопровождаемого образованием кольцевой структуры, которая продолжает существовать в виде внехромосомного элемента (двойной минихромосомы (double-minutes – DM)), способного к автономной репликации. Предполагают, что DM является промежуточным продуктом амплификации, образующимся в условиях низкого давления селектирующего фактора окружающей среды. На более поздних стадиях амплифицирующаяся ДНК после повторной интеграции в геном обнаруживается преимущественно в составе удлиненного участка хромосомы (extended chromosomal region – ECR), иначе называемого гомогенно окрашивающейся областью (homogeneously staining region – HSR). На этой стадии амплифицированный участок генома клеток, а следовательно, и их фенотип становятся более стабильными. Для биотехнологических целей в системе амплификации рекомбинантных генов в качестве селектируемого маркера часто используют ген дигидрофолатредуктазы (ДГФР), интегрированный в экспрессирующий вектор. ДГФР катализирует превращение фолиевой кислоты в тетрагидрофолат, необходимый для синтеза эукариотическими клетками глицина, тимидинмонофосфата и пуриновых оснований. В этой связи клетки CHO, в которых ген ДГФР инактивирован под действием мутации, не растут на средах без нуклеозидов и приобретают эту способность после трансфекции геном ДГФР. Растущие трансфектанты далее отбираются по признаку амплификации гена ДГФР на фоне увеличивающихся концентраций метотрексата – ингибитора ДГФР, в питательной среде, так как увеличение числа копий гена будет придавать клеткам устойчивость к ингибитору в больших концентрациях. После проведения множественных раундов селекции получают популяцию клеток, содержащих до нескольких сотен копий гена ДГФР. 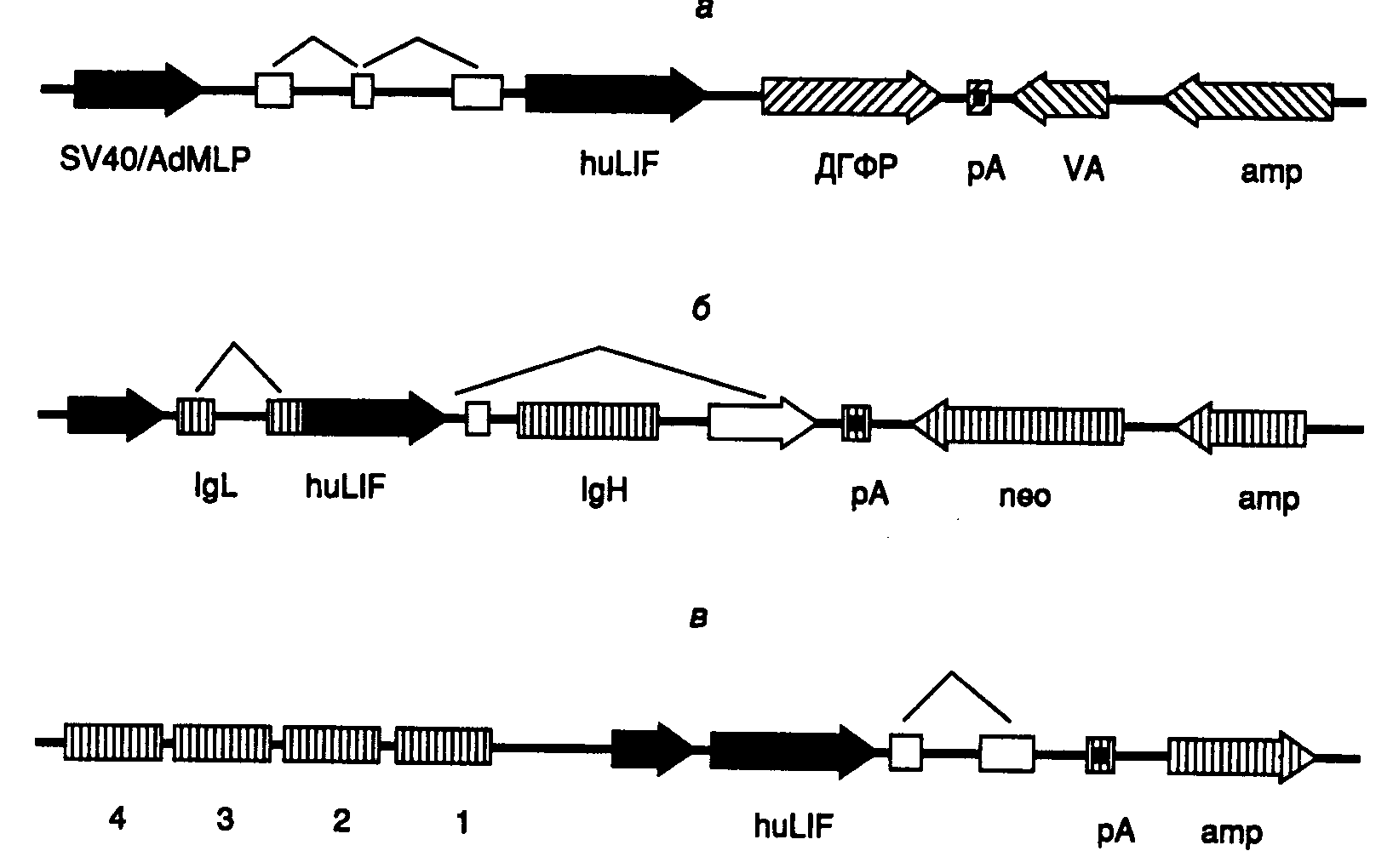 Рис. II.13. Кассеты генов в экспрессирующих векторах, использованные для получения фактора, ингибирующего лейкозы, в культурах клеток а – кДНК huLIF (черные стрелки), включающая лидерные последовательности (незаштрихованные прямоугольники), была интегрирована в экспрессирующий вектор pXMT3 под контроль гибридного промотора, расположенного между точкой начала репликации вируса SV40 и основным поздним промотором аденовируса (SV40/AdMLP), ДГФР, amp, neo – гены дигидрофолатредуктазы и устойчивости к ампициллину и неомицину соответственно, VA – трансляционный энхансер; pA – сайт полиаденилирования; б – вектор pSV2neo-Ig-huLIF, сконструированный на основе транскрипционных и трансляционных регуляторных элементов генов тяжелой (IgH) и легкой (IgL) цепей иммуноглобулинов. Транскрипция кассеты контролируется мышиным промотором гена IgL и энхансером гена IgH. Транскрипт, содержащий последовательность huLIF, в результате сплайсинга объединяется с экзоном 2 гена IgL, кодирующего лидерную последовательность, которая обеспечивает секрецию зрелого huLIF клетками; в – вектор pGES-LIF, прямоугольники 1–4 – регуляторная область кластера глобиновых генов, короткая черная стрелка – промотор -глобинового гена, ломаной линией отмечены последовательности, удаляемые сплайсингом. Во всех случаях стрелки указывают направление транскрипции Размер амплифицированного геномного локуса значительно больше размера селектируемого гена и может составлять несколько сотен тысяч пар оснований. Трансфекция клеток геном ДГФР и исследуемым рекомбинантным геном, заключенными в один экспрессирующий вектор, сопровождаемая их интеграцией в геном, будет приводить к их совместной амплификации на фоне метотрексата и усилению синтеза требуемого рекомбинантного белка, как следствие увеличения дозы его гена. Данные о возможности сохранения высокого уровня экспрессии гена в отсутствие селектирующего агента, т.е. генетической стабильности продуцентов, противоречивы. Результат зависит от генотипа конкретных клонов клеток, в том числе и локализации сайта интеграции экспрессирующего вектора в геном клеток CHO. Эффективным экспрессирующим вектором, используемым в описываемой системе, является двухцистронная молекула ДНК, в которой исследуемый рекомбинантный ген находится под контролем сильного промотора перед геном ДГФР, что обеспечивает высокое сопряжение синтеза рекомбинантного белка с уровнем устойчивости клеток к метотрексату (рис. II.13,а). 7.4.2.Клетки мышиной миеломы (линия Sp2/0)Клетки миеломы находят широкое применение для получения гибридом – линий клеток, производящих моноклональные антитела определенной специфичности. Поскольку эти клетки осуществляют эффективную секрецию рекомбинантных белков, они хорошо изучены в отношении экспрессии в них соответствующих рекомбинантных генов, введенных с помощью трансфекции. Кроме того, такие клетки способны расти в суспензионной культуре (без прикрепления к поверхности субстрата), что облегчает их культивирование при необходимости крупномасштабной наработки. Линия клеток мышиной миеломы Sp2/0 Ag14, утративших способность синтезировать или секретировать иммуноглобулины, часто используется для производства рекомбинантных белков. Клетки этой линии легко трансфецируемы рекомбинантными ДНК и могут выращиваться в больших количествах на питательной среде без дорогостоящей сыворотки. Клетки Sp2/0 экспрессируют эндогенный ген ДГФР, что делает невозможным их прямое применение для амплификации рекомбинантных ДНК с использованием метотрексата. Это затруднение обычно преодолевают путем введения в экспрессирующий вектор мутантного гена ДГФР, в котором точковая мутация, сопровождаемая заменой TG, приводит к замещению LeuArg в положении 22 полипептидной цепи ДГФР, следствием чего является понижение сродства фермента к метотрексату в 270 раз. Благодаря этому после трансфекции может быть проведен отбор клеток, содержащих мутантный ген ДГФР вместо нормального, путем постепенного увеличения концентрации метотрексата в среде. При умеренных концентрациях ингибитора эндогенный ген ДГФР полностью инактивируется, тогда как мутантный ген остается активным и может быть амплифицирован в результате описанного выше механизма после повышения концентрации метотрексата в среде. Однако уровень амплификации гена ДГФР и ассоциированного с ним рекомбинантного гена в такой системе как правило ниже, чем в обычной, так как меньшее число внутриклеточных копий мутантного гена требуется для обеспечения необходимого уровня устойчивости клеток к ингибитору. Кроме того, было показано, что высокий уровень внутриклеточной экспрессии рекомбинантных генов, введенных в клетки Sp2/0 с помощью трансфекции, может обеспечиваться после инкубации с метотрексатом за счет увеличения скорости их транскрипции, а не в результате их амплификации. Такой результат может быть получен в результате введения множественных копий гена ДГФР при трансфекции. Структура одного из векторов, пригодного для использования с миеломными клетками Sp2/0, представлена на рис. II.13,б). Транскрипция трансгенов в этом векторе обеспечивается промоторно–энхансерными элементами гена иммуноглобулина с участием эндогенных регуляторных факторов клеток миеломы. 7.4.3.Клетки селезенки мышей (линия MEL)Обе системы экспрессии, описанные выше, базируются на амплификации трансгенов, обеспечивающей высокий уровень внутриклеточного синтеза кодируемых ими рекомбинантных белков в отобранных клонах клеток. Тем не менее, в природе уникальные гены (т.е. присутствующие в виде одной копии на гаплоидный геном) могут экспрессироваться с очень высокой эффективностью. Примером этого являются, например иммуноглобулины, секретируемые плазматическими клетками, или глобины, синтезируемые в больших количествах клетками эритроидного ряда. Высокий уровень транскрипции глобинового локуса становится возможным не только благодаря функционированию эффективных промотора и энхансера, но и в результате присутствия так называемой доминантной регуляторной области (dominant control region – DCR), изменяющей пространственную структуру хроматина всего локуса. DCR расположена за |