|
|
Рецензенты кандидат химических наук В. Г. Коробко доктор биологических наук В. А. Гвоздев Патрушев Л. И
7.2.5.Интегрирующие и челночные (бинарные) векторы
Векторы, пригодные для клонирования ДНК в бактериях, отличающихся от E. coli, должны обладать всеми характерными чертами, которые были отмечены выше. От только что рассмотренных они отличаются главным образом тем, что содержат природные или искусственные генетические элементы, функционирующие в новых клетках-хозяевах.
Интегрирующие векторы грамположительной бактерииBacillus subtilis. B. subtilis, как и E. coli, является излюбленным объектом генной инженерии. Это связано с тем, что B. subtilis – непатогенный микроорганизм, многие штаммы которого широко используются в микробиологической промышленности для производства биологически активных соединений и пищевых веществ. В отличие от E. coli B. subtilis способна секретировать белки и пептиды, что облегчает их очистку и дальнейшее использование. Большинство векторов для клонирования ДНК в клетках B. subtilis создано на основе плазмид других видов бацилл, а также Staphylococcus aureus и бактерий рода Streptococcus.
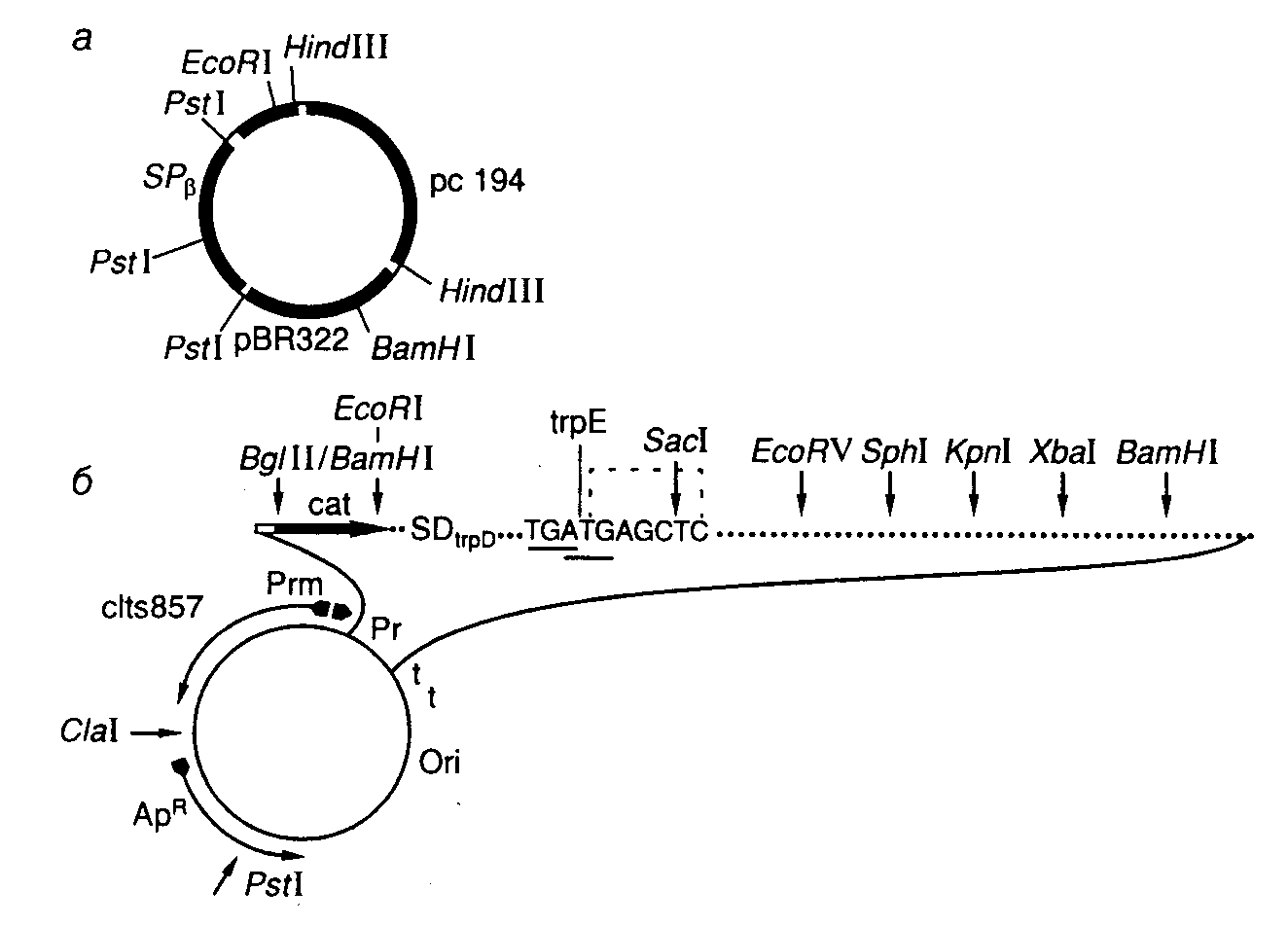
Рис. II.11. Челночный вектор pFH7 (а) и экспрессирующий вектор pPR-TGATG-1 (б)
Обозначено расположение генов и уникальных сайтов рестрикции
Основным свойством интегрирующих векторов является их способность стабильно встраиваться в геном клетки-хозяина. Это становится возможным благодаря наличию в таких векторах последовательностей нуклеотидов, гомологичных последовательностям геномной ДНК. В результате функционирования общей системы рекомбинации происходит объединение хромосомной и плазмидной ДНК интегрирующего вектора, которое приводит к стабильному включению всей векторной плазмиды в хромосому. Примером такого интегрирующего вектора служит плазмида pFH7 B. subtilis (рис. II.11,а). Векторная плазмида содержит фрагмент ДНК умеренного бактериофага SP и после попадания в клетки B. subtilis, лизогенные по данному бактериофагу, эффективно интегрируется в профаг. Поскольку плазмида содержит ген устойчивости к хлорамфениколу cat, клетки приобретают этот признак. Индукция профага приводит к образованию фаговых частиц, трансдуцирующих такую плазмиду и ассоциированный с ней признак устойчивости к хлорамфениколу. Интеграция плазмиды SP в бактериальную хромосому происходит по механизму гомологичной рекомбинации с участием гена recE.
Способность к интеграции в бактериальную хромосому обнаруживают и другие плазмиды, содержащие фрагменты хромосомной ДНК клеток-хозяев, что продемонстрировано, в частности, для плазмид E. coli и Streptococcus pneumoniae.
Интегрирующие векторные системы, в которых используется тот же принцип гомологичной рекомбинации, разработаны и для эукариотических клеток, включая клетки животных и растений. В конце концов, такие работы привели к развитию целого направления исследований по созданию трансгенных животных и растений, стабильно наследующих и экспрессирующих гены, искусственно введенные в их геном. О некоторых важных следствиях этого направления исследований, включая генотерапию, речь пойдет в главе 10.
Наличие феномена гомологичной рекомбинации между хромосомной ДНК клетки-хозяина и векторной ДНК, содержащей гомологичные хромосомной ДНК последовательности нуклеотидов, приходится учитывать при получении соответствующих генно-инженерных конструкций. Такая неконтролируемая рекомбинация в большинстве случаев нежелательна, так как может приводить к потере или структурным перестройкам клонированных фрагментов ДНК. Для того чтобы свести последствия этого явления к минимуму, используют специальные штаммы клеток-хозяев, в которых общая система рекомбинации не функционирует, например вследствие мутационной инактивации гена recA E. coli или recE B. subtilis.
Челночные векторы. Интегрирующая плазмида pFH7 (см. рис. II.11,а) дает возможность проиллюстрировать еще один важный принцип, широко используемый при конструировании векторных систем в генной инженерии. Эта плазмида получена путем объединения двух репликонов, один из которых берет начало от плазмиды pC194 B. subtilis, а другой – от плазмиды pBR322 E. coli, что дает возможность вектору существовать и стабильно реплицироваться как в клетках E. coli, так и B. subtilis. Такие векторы, способные реплицироваться в клетках-хозяевах разных биологических видов, называют челночными, или бинарными векторами.
Принципы конструирования и функционирования челночных векторов одинаковы, они должны включать в себя репликоны тех генетических систем, в которых будет происходить репликация челночного вектора. При этом используются области начала репликации генетических элементов, которые автономно существуют во внехромосомном состоянии в природных условиях. Так, обсуждавшийся выше интегрирующий вектор pFH7 B. subtilis обладает свойствами челночного вектора, поскольку для его конструирования использованы репликоны двух видов бактерий. Более эффектными примерами челночных векторов являются плазмидные ДНК, способные реплицироваться в клетках высших (животных и растений) и низших организмов. Необходимость использования челночных векторов в генной инженерии связана с тем, что наработку в препаративном количестве векторной ДНК для проведения генно-инженерных манипуляций удобнее проводить в бактериальных клетках, тогда как получение биологически активных продуктов клонированных генов высших организмов во многих случаях возможно только в клетках своего или близкого вида, в которых эти гены экспрессируются в природных условиях, т.е. в своем обычном генетическом окружении (подробнее см. раздел 7.6).
7.2.6.Конструирование экспрессирующих векторов и их функционирование
Первая часть книги была посвящена описанию механизмов, обеспечивающих высокоэффективную и высокоспецифическую экспрессию генов. Такого рода информацию успешно используют в настоящее время для эффективной экспрессии рекомбинантных генов в гомологичном или гетерологичном генетическом окружении. После рассмотрения основных принципов конструирования векторов для клонирования ДНК можно перейти к обсуждению проблемы экспрессии клонированных генов в искусственных генетических системах. Именно экспрессия клонированных генов является одной из основных задач генной инженерии. Действительно, функциональную роль отдельных генов и их частей в живом организме можно понять и оценить лишь по их экспрессии, т.е. по фенотипическому проявлению их потенциальных биологических возможностей. Для получения полноценной экспрессии клонированных генов используют экспрессирующие векторные системы, принципы конструирования которых в настоящее время хорошо разработаны.
Напомним, что экспрессия генетической информации, заключенной в генах живых организмов, в соответствии с основным постулатом молекулярной биологии осуществляется путем ее переноса от нуклеиновых кислот к белкам по схеме: ДНК РНК белок. Любой полноценный в функциональном отношении ген, в том числе и рекомбинантный, состоит из двух основных частей – регуляторной и структурной. Генетический код, с помощью которого последовательность нуклеотидов в кодирующей структурной части гена определяет последовательность аминокислот в кодируемом этим геном белке, за немногими исключениями, универсален для всех живых организмов. Это дает принципиальную возможность получения полноценной экспрессии самых разнообразных осмысленных последовательностей нуклеотидов, в том числе и искусственно синтезированных, в любом природном генетическом окружении. Таким образом, универсальность генетического кода является прочной основой для создания функционально активных генно-инженерных конструкций.
Регуляторная часть генов распознается соответствующими ферментными системами организма и обеспечивает упорядоченную экспрессию его структурной части. При этом регуляторная часть гена, как правило, высокоспецифична в отношении своих природных генетических эффекторов (РНК-полимераз, рибосом, факторов транскрипции и трансляции, белковых факторов сплайсинга, ферментов, осуществляющих посттрансляционные модификации полипептидов, и т.п.), и чаще всего она не может эффективно функционировать в гетерологичном генетическом окружении. Очевидно, что при конструировании высокоэффективных экспрессирующих векторов необходимо, прежде всего, учитывать особенности структуры регуляторной части рекомбинантного гена, исходя из того, в каких генетических условиях клонированный ген предполагается экспрессировать. Однако не только регуляторные последовательности генов являются препятствием для высокоэффективной экспрессии чужеродных рекомбинантных генов. Структурные части генов про- и эукариот фундаментально отличаются друг от друга по наличию у последних внутри генов многочисленных и протяженных некодирующих последовательностей нуклеотидов – интронов. Следовательно, гены эукариот не могут эффективно экспрессироваться в клетках прокариот, поскольку у прокариот отсутствуют соответствующие системы сплайсинга. Кроме того, у предшественников эукариотических мРНК не может осуществиться в бактериальных клетках и правильный процессинг 3’- и 5’-концевых некодирующих последовательностей. Даже такой беглый взгляд на проблему экспрессии рекомбинантных генов в чужеродном генетическом окружении позволяет увидеть многочисленные препятствия, лежащие на пути решения этой проблемы. Ниже будут рассмотрены факторы, которые необходимо учитывать при конструировании экспрессирующих векторов.
Факторы, оказывающие влияние на эффективность экспрессии рекомбинантных генов: транскрипция. Для того чтобы в клетках E. coli или любого другого организма произошла экспрессия клонированного гена, этот ген должен быть транскрибирован РНК-полимеразой, а образовавшаяся РНК транслирована рибосомами с образованием нативного белкового продукта. Следовательно, для эффективной экспрессии в клетках E. coli любой ген должен находиться под контролем регуляторных элементов данного микроорганизма. Особенности структуры и функционирования регуляторных элементов генов про- и эукариот были подробно рассмотрены в первой части книги. Знание основных принципов строения и функционирования промоторов позволяет конструировать новые промоторы, которые по своей эффективности могут превосходить природные. Примером такого промотора, используемого в некоторых экспрессирующих векторах, является промотор Taq, который в своем составе объединяет последовательности промоторов лактозного и триптофанового оперонов E. coli.
Базальный уровень транскрипции рекомбинантных генов гарантируется небольшим набором упорядоченных регуляторных последовательностей. Например, у E. coli это происходит при наличии перед структурной частью клонированного гена последовательностей в окрестностях нуклеотидов +1, 10 и –35.
-35 -10 +1
a.....tcTTGACat..t........t.tg.TAtAat........cat
Однако даже у E. coli транскрипция большинства генов носит регулируемый характер, что обеспечивается системой белков-репрессоров и активаторов, а также низкомолекулярных эффекторов, которые главным образом влияют на взаимодействие РНК-полимеразы с транскрибируемыми генами. Необходимость регулирования экспрессии генов в генной инженерии диктуется, прежде всего, тем, что сверхэкспрессия клонированных генов в клетках-продуцентах рекомбинантных белков при использовании сильных конститутивных промоторов может отрицательно сказываться на их жизнеспособности, поскольку промежуточные и конечные продукты экспрессии часто обладают цитотоксическим действием. В этом случае сверхэкспрессия клонированных генов сопровождается снижением скорости или даже полной остановкой роста бактериальных или иных клеток. Для предотвращения такого рода эффектов в генной инженерии и биотехнологии используют генно-инженерные конструкции, в которых экспрессия клонированных генов в значительной степени подавлена (репрессирована) на ранних фазах роста культуры рекомбинантных клеток и может быть дерепрессирована в любой нужный момент времени.
В качестве примера рассмотрим две системы регулируемой экспрессии рекомбинантных генов в клетках E. coli, которые получили широкое распространение. В одном случае для контроля экспрессии клонированных генов используют хорошо изученную систему регуляторных элементов лактозного оперона, в другом – систему промотор–репрессор–оператор фага . Принцип регуляции экспрессии рекомбинантных генов в обоих случаях один и тот же. В векторные молекулы вводятся регуляторные последовательности фага или lac-оперона, за которыми следуют последовательности нуклеотидов полилинкера с несколькими уникальными сайтами рестрикции, по которым проводится клонирование исследуемого гена. Одновременно в тот же вектор вводится ген-регулятор, кодирующий белок-репрессор. В отсутствие индуцирующего воздействия молекулы репрессора, ген которого экспрессируется конститутивно, связываются с оператором, тем самым препятствуя взаимодействию РНК-полимеразы с промотором, под контролем которого находится клонированный рекомбинантный ген. Это приводит к резкому снижению уровня транскрипции клонированного гена, низкому внутриклеточному содержанию соответствующей мРНК и белкового продукта ее трансляции.
Существенным различием двух систем регулируемой экспрессии является способ их индукции. В случае регуляторной системы lac-оперона чаще всего используют синтетический аналог природного индуктора (алло-лактозы) – изопропил–-D-тиогалактопиранозид (IPTG). Индуктор связывается с молекулой репрессора и нарушает его взаимодействие с операторным участком ДНК. Область lac-промотора становится доступной для РНК-полимеразы, которая с высокой эффективностью начинает транскрибировать гены, расположенные вслед за промотором. В отличие от этого в системе репрессии-дерепрессии фага используют ген репрессора cI, содержащий температурочувствительную мутацию. Наличие такой мутации приводит к тому, что мутантный белок-репрессор сохраняет свою нативную конформацию, а следовательно, и способность подавлять синтез РНК только при пермиссивной температуре (как правило, 28–30о). При повышении температуры окружающей среды до 42о происходят инактивация белка-репрессора и дерепрессия генов, расположенных под контролем промоторов PL или PR фага .
Использование системы репрессии–дерепрессии, чувствительной к температуре, более технологично по сравнению с химической индукцией по двум обстоятельствам: во-первых, это менее дорогой способ индукции; во-вторых, для системы фага характерна более высокая степень репрессии генов, находящихся под ее контролем, и соответственно более низкий базальный уровень их экспрессии. Такой фактор становится решающим при клонировании и получении экспрессии генов, белковые продукты которых токсичны для клетки-хозяина. Однако несмотря на технологичность системы репрессии–дерепрессии, чувствительной к температуре, сверхпродукция рекомбинантных белков, особенно эукариотических, часто сопровождается внутриклеточным образованием их нерастворимых агрегатов, что в ряде случаев полностью отсутствует при использовании для выращивания рекомбинантных клеток температур ниже 30о (подробнее см. ниже). Эти эффекты накладывают ограничения на использование векторов с температурочувствительным контролем экспрессии для продукции эукариотических и в меньшей степени прокариотических белков в бактериальных клетках.
Для обеспечения регулируемой тканеспецифической экспрессии рекомбинантных генов в соматических клетках животных и растений в составе векторов используют энхансеры, которые избирательно стимулируют транскрипцию в соответствующих тканях и не оказывают такого действия на гены в тканях, клетки которых не экспрессируют необходимые регуляторные белки. Кроме того, популярным становится введение в экспрессирующие эукариотические векторы пограничных последовательностей нуклеотидов, фланкирующих клонируемые гены, которые помогают обеспечивать экспрессию рекомбинантных генов, сводя к минимуму эффект их положения в хромосомах соматических клеток. О механизмах этих явлений подробно говорилось в первой части книги.
В настоящее время, по-видимому, не существует систем репрессии клонируемых генов, которые бы полностью блокировали их транскрипцию. Это создает определенные проблемы при клонировании генов белков, токсичных для клеток-хозяев, таких как колицины, эндо- и экзонуклеазы, протеиназы и т.п.
Наряду с промоторами векторы, экспрессирующие рекомбинантные гены в клетках E. coli, должны содержать и гомологичные терминаторы транскрипции. Необходимость в этом вызвана тем, что даже нетранслируемые избыточные 3’-концевые последовательности нуклеотидов мРНК, транскрибированной с рекомбинантного гена, отрицательно сказываются на эффективности ее трансляции, что, по-видимому, связано с участием таких последовательностей в образовании сложной третичной структуры рекомбинантных мРНК.
Факторы, влияющие на эффективность экспрессии рекомбинантных генов: трансляция. Как следует из изложенного выше, эффективность экспрессии клонированных генов в бактериальных и эукариотических клетках во многом зависит от эффективности их транскрипции, т.е. от уровня внутриклеточного содержания соответствующих мРНК. Однако эффективность экспрессии рекомбинантных генов не в меньшей степени зависит и от эффективности трансляции таких мРНК рибосомами. При работе с рекомбинантными ДНК чаще всего приходится иметь дело с гетерологичными системами, в которых клонированные экспрессируемые ДНК чужеродны по отношению к генетическому окружению и ферментативным системам бактериальных или эукариотических клеток-хозяев. В первой части книги в качестве примеров были рассмотрены некоторые факторы, играющие большую роль в регуляции экспрессии генов на посттранскрипционном уровне. Такие факторы вносят свой вклад и в эффективность экспрессии рекомбинантных генов. Однако в этом случае необходимо всегда иметь в виду уровень соответствия регуляторных факторов клеток-хозяев регуляторным последовательностям мРНК, которые являются транскриптами рекомбинантных генов. В частности, на эффективность трансляции чужеродных мРНК в бактериальных клетках оказывает сильное влияние первичная структура небольшого участка 5’-концевой части мРНК перед инициирующим AUG-кодоном, частично комплементарная 3’-концевой части 16S рРНК. Этот участок мРНК называют последовательностью Шайна–Дальгарно, или SD-последовательностью.
SD-последовательность мРНК служит сигналом для взаимодействия рибосом с мРНК и обеспечивает инициацию трансляции с правильного AUG-кодона. Она представляет собой короткую последовательность нуклеотидов, обогащенную пуриновыми основаниями, и расположена за 4–9 нуклеотидов перед AUG-кодоном:
16S рРНК HOAUUCCUCCACUAGG………………5’
lacZ-мРНК UUCACACAGGAAACAGCUAUGACCAUGAUU………3’
(в приведенном примере прописными буквами у lacZ-мРНК изображена SD-последовательность нуклеотидов, комплементарная соответствующей пиримидиновой последовательности 16S рРНК, и подчеркнут инициирующий кодон). Различия в эффективности распознавания такого сайта на мРНК рибосомами приводят к различиям в эффективности экспрессии соответствующих генов. Достаточно наличия в SD-сайте трех нуклеотидов, комплементарных соответствующему участку 16S РНК, чтобы рибосомы приобретали способность связывать эту мРНК. Помимо SD-последовательности в мРНК, по-видимому, имеются и другие участки, необходимые для эффективной инициации трансляции рибосомами, так как на основании только первичной структуры мРНК в настоящее время невозможно предсказать эффективность сайта связывания рибосом. Более того, сайты инициации трансляции на мРНК, обладающие протяженной гомологией с 16S рРНК, являются малоэффективными, возможно, из-за слишком прочного связывания рибосом с мРНК.
Существенным фактором, оказывающим влияние на эффективность трансляции мРНК рибосомами, может быть вторичная структура мРНК в окрестностях инициирующего кодона. Из всего вышеизложенного следует, что некодирующая 5’-концевая часть мРНК играет важную роль в эффективности ее трансляции рибосомами, и, по крайней мере, эти особенности их трансляции необходимо учитывать при конструировании векторов для экспрессии рекомбинантных генов в клетках E. coli. Современные экспрессирующие векторы построены таким образом, что промотор, с которого начинается транскрипция клонированного гена, а также следующая за ним последовательность нуклеотидов, включающая инициирующий ATG-кодон, являются частью генов, эффективно экспрессирующихся в клетках E. coli. Сразу вслед за ATG-кодоном вектора располагают уникальные сайты рестрикции, по которым проводят встраивание кодирующей части клонируемого гена, экспрессию которого хотят получить. В том случае, если сайт рестрикции, по которому производится клонирование, расположен в начале кодирующей части природного гена, в результате транскрипции такой рекомбинантной молекулы образуется химерная мРНК. 5’-Концевая часть этой мРНК от промотора до сайта рестрикции для клонирования соответствует последовательности нуклеотидов вектора и кодирует несколько аминокислот природного гена, регуляторные последовательности которого использовали для конструирования экспрессирующего вектора. Протяженная же 3’-концевая часть такой мРНК будет соответствовать клонированной последовательности нуклеотидов. Если соединение этих двух частей мРНК произошло в одной рамке считывания, то в результате ее трансляции будет образовываться химерный полипептид, N-концевая часть которого кодируется вектором, а С-концевая – клонированным фрагментом ДНК. Примерами таких экспрессирующих векторов могут быть плазмиды pUC18 и pUC19 (см. рис. II.5,б), продукты экспрессии которых представляют собой химерные белки, а N-концевая часть содержит N-концевые аминокислотные остатки -галактозидазы E. coli.
Превосходным вектором, прекрасно зарекомендовавшим себя в работе, является экспрессирующий вектор pPR-TGATG-1, сконструированный в лаборатории С.В. Машко (г. Москва). Вектор функционирует по тому же самому принципу. В нем структурная часть экспрессируемого гена соединяется через уникальный сайт рестрикции SacI с инициирующим кодоном ATG (см. рис. II.11,б). В результате образуется гибридный оперон, транскрибируемый с промотора -PR, дистальный цистрон которого представлен клонируемым геном, а цистрон, проксимальный промотору PR, содержит гибридный ген cro’-cat’-trpE, состоящий из слитых частей генов cro фага , хлорамфениколацетилтрансферазы cat и trpE E. coli. 3’-Концевая часть цистрона содержит область реинициации трансляции, SD-последовательность гена trpD E. coli и перекрывающиеся кодоны терминации и инициации трансляции (TGATG). В этой последовательности 3’-концевой нуклеотид G является первым нуклеотидом уникального сайта рестрикции SacI (GAGCTC). В процессе синтеза белка, направляемого вектором, образуется короткий гибридный пептид Cro’-Cat’-TrpE длиной
120 аминокислот. Сразу же после терминации его синтеза с высокой эффективностью происходит реинициация синтеза полипептида, кодируемого клонированным геном, на перекрывающемся с кодоном TGA ATG-кодоне. Поскольку промотор PR в этом векторе контролируется температурочувствительным репрессором фага cIts857, экспрессия клонированного гена индуцируется повышением температуры окружающей среды до 42о и сопровождается инактивацией репрессора.
Приведенные примеры четко демонстрируют роль двух основных факторов, оказывающих влияние на эффективность экспрессии чужеродных клонированных генов, находящихся в составе экспрессирующих векторов в бактериальных клетках. Этими факторами являются, во-первых, оптимальная транскрипция клонированных генов и, во-вторых, эффективная трансляция транскрибированной мРНК. В обоих случаях наилучшие результаты могут быть получены при объединении в составе вектора структурной части клонированного чужеродного гена с генетическими элементами, регулирующими транскрипцию и трансляцию (промоторы, SD-последовательности и т.п.) бактериальных клеток-хозяев или их вирусов. Знание главных принципов, лежащих в основе регуляции транскрипции и трансляции, позволяет конструировать новые регуляторные последовательности (например промотор Taq) путем объединения известных регуляторных элементов или использования искусственно синтезированных последовательностей нуклеотидов, представляющих собой усредненные канонические регуляторные последовательности, учитывающие особенности большого числа природных регуляторных элементов. Однако этими важными факторами не исчерпывается возможность оптимизации экспрессии рекомбинантных генов в чужеродном генетическом окружении.
Среди других факторов, оказывающих влияние на эффективность трансляции, следует упомянуть частоту встречаемости кодонов при кодировании белков в структурных частях разных генов. В настоящее время установлено, что использование синонимических кодонов (кодирующих одну и ту же аминокислоту) вырожденного генетического кода не случайно и отражает количественную представленность отдельных изоакцепторных тРНК в клетках организма. С другой стороны, частота использования кодонов в разных генах одного и того же организма является эффективным фактором, регулирующим уровень экспрессии этих генов в процессе трансляции. Чем реже тот или иной кодон используется для кодирования аминокислоты в данном организме, тем с меньшей скоростью он будет транслироваться рибосомами вследствие низкой внутриклеточной концентрации тРНК, узнающей такой кодон. Так, в группе белков дрожжей с высоким уровнем экспрессии 96% аминокислот кодируются только 25 из 61 возможного кодона. Замена этих 25 предпочтительных кодонов в 5’-концевых частях генов белков с высоким уровнем экспрессии на редко встречающиеся кодоны снижает уровень синтеза как белков, так и их мРНК.
Кроме того, на эффективность трансляции кодона оказывает влияние и контекст кодона, т.е. соседние, окружающие кодон последовательности нуклеотидов. Этот феномен предпочтительного использования одних кодонов перед другими необходимо учитывать в опытах по оптимизации экспрессии чужеродного генетического материала в гетерологичном генетическом окружении, поскольку несоответствие в частотах использования кодонов у организмов разных видов может служить серьезным препятствием для получения эффективной экспрессии клонируемых генов. Снижение уровня экспрессии под влиянием подобных факторов, в частности, наблюдали при попытках экспрессии генов растений в клетках дрожжей, а также в других гетерологичных системах. Таким образом, эффекты, возникающие в связи с различной частотой использования кодонов, необходимо учитывать при подборе генетических систем для эффективной экспрессии клонированных чужеродных генов. Этот фактор может оказаться одним из решающих при получении полностью синтетических генов, например генов пептидных гормонов, для использования их в биотехнологии, а также при планировании мутационных замен нуклеотидов в белковой инженерии.
Факторы, влияющие на эффективность экспрессии рекомбинантных генов: стабильность рекомбинантных белков и РНК. Важным фактором, определяющим уровень экспрессии рекомбинантных генов в реципиентных клетках, является стабильность кодируемых этими генами мРНК и белков. В первой части книги подробно рассматривалось значение таких факторов в регуляции экспрессии генов. Не менее важную роль эти феномены играют и в контроле уровня экспрессии рекомбинантных генов в искусственных генетических системах.
Как было упомянуто выше, замены кодонов в мРНК дрожжей на синонимические приводят не только к снижению уровня трансляции измененных мРНК в клетках дрожжей, но и сопровождаются уменьшением стабильности самих мРНК. Требование достаточного уровня стабильности мРНК является одним из факторов, определяющих давление отбора на использование конкретных синонимических кодонов для кодирования белков. Важную роль во внутриклеточной стабильности мРНК играют элементы ее вторичной структуры. Так, образование шпилек на 3’-конце мРНК препятствует ее деградации под действием бактериальных экзонуклеаз, РНКазы II и полинуклеотидфосфорилазы. Поскольку изменение первичной структуры с учетом возможной вторичной структуры без изменения функциональных свойств представляется в настоящее время сложной задачей, проблема внутриклеточной стабильности рекомбинантных РНК решается главным образом путем введения мутаций в гены экзо- и эндонуклеаз бактериальной клетки-хозяина. Так называемые безРНКазные штаммы E. coli находят применение в качестве реципиентов рекомбинантных плазмид в генной инженерии. При этом решается двойная задача: с одной стороны, в таких клетках повышается стабильность транскриптов рекомбинантных генов, а с другой – пониженный внутриклеточный уровень определенных РНКаз способствует более эффективной очистке рекомбинантных белков, так как для многих препаратов ферментов даже незначительные примеси нуклеазной активности бывают нежелательны.
Проблема внутриклеточной стабильности рекомбинантных белков больше связана с деградацией небольших пептидов, поскольку крупные нативные белки более стабильны в бактериальных клетках. Один из подходов, позволяющих стабилизировать короткие чужеродные пептиды в клетках E. coli, – включение требуемого пептида в состав гибридного белка. В этом случае последовательность нуклеотидов, кодирующую гибридный белок, соединяют в составе экспрессирующего вектора в одной рамке считывания с бактериальным геномом, кодирующим белок (например геном -галактозидазы). Образующийся в результате экспрессии такого рекомбинантного гена гибридный белок в своем составе содержит в N- или С-концевой части требуемый пептид, защищенный основным белком-носителем от протеолитической деградации. Для отделения пептида от белка-носителя их соединяют друг с другом последовательностью аминокислот, по которой можно провести специфическое расщепление полипептидной цепи. В том случае, если пептиды не содержат метионина, соединение белка-носителя и пептида осуществляют через эту аминокислоту, и отщепление пептида производят с помощью бромистого циана. Такой подход был использован, например для получения рекомбинантного соматостатина, а также A- и B-цепей инсулина.
Для разделения рекомбинантного пептида и белка-носителя можно использовать и протеолитические ферменты. Для этого требуемый пептид соединяют с белком-носителем с помощью последовательности аминокислот, специфически расщепляемой протеолитическим ферментом, например пептидазой клостридий (клострипаином) в случае получения кальцитонина или трипсином для получения -эндорфина. Второй подход, позволяющий защищать рекомбинантные белки от внутриклеточного протеолиза, – выведение их в периплазматическое пространство или окружающую среду после завершения биосинтеза. При таком подходе рекомбинантные полипептиды соединяют с сигнальными последовательностями аминокислот белков (-лактамаза, OmpA, OmpF, PhoA), секретируемых во внешнюю среду и периплазматическое пространство. Подобным путем были получены, например проинсулин, гормон роста человека и -эндорфин.
Особенности биосинтеза рекомбинантных эукариотических белков в бактериальных клетках: тельца включения. Экспрессия эукариотических генов в бактериальных клетках связана еще с одной проблемой: большая часть рекомбинантных эукариотических белков, а также некоторые прокариотические белки при очень высоком уровне биосинтеза переходят в нерастворимое состояние, образуя в клетках так называемые тельца включения. Тельца включения представляют собой частицы, состоящие главным образом из агрегатов рекомбинантного белка и ряда бактериальных белков. Рекомбинантные белки в тельцах включения находятся, как правило, в денатурированном неактивном состоянии, и для их получения в растворимом виде приходится применять мощные денатурирующие агенты, такие как мочевина, гуанидинхлорид и детергенты. Солюбилизированный в этих жестких условиях рекомбинантный белок для восстановления нативной конформации требует ренатурации, в процессе обработки денатурирующими агентами белки могут изменять свою структуру, а при концентрировании из разбавленных растворов, в которых проводится ренатурация, часто вновь выпадают в осадок.
В чем же причина такого аномального поведения рекомбинантных эукариотических белков в бактериальных клетках? Для поддержания белков в растворимом состоянии в клетках эукариот реализуются три основных механизма: компартментализация продуктов трансляции, белок-белковые взаимодействия и посттрансляционные модификации. Образующиеся в результате трансляции мРНК рибосомами в эндоплазматическом ретикулуме полипептидные цепи не распределяются хаотически в цитоплазме эукариотических клеток, но последовательно переходят через ряд компартментов, где претерпевают посттрансляционные модификации. При этом внутренние условия отдельных компартментов могут существенно различаться. Так, молекулы инсулина накапливаются in vivo в секреторных гранулах, pH в которых составляет 4,5–5,5. В то же время pH цитоплазмы бактериальных клеток приближается к 7,8, в этих условиях инсулин лишь слабо растворим. Многие гидрофобные эукариотические белки не существуют в растворимой форме в отсутствие фосфолипидов мембран. Белки молока являются интегральной частью мицелл, стабилизированных ионами Ca2+.
Посттрансляционные модификации белков, в частности фосфорилирование, гидроксилирование, гликозилирование и ограниченный протеолиз, происходящие в определенных компартментах эукариотических клеток, также оказывают большое влияние на растворимость белков, их стабильность и биологические функции. Это особенно относится к секретируемым белкам эукариот. Поскольку в бактериальных клетках соответствующие системы посттрансляционных модификаций белков отсутствуют, в них не могут быть получены биологически полноценные рекомбинантные эукариотические полипептиды, что накладывает ограничения на использование бактерий в биотехнологии.
Наконец, третьим ключевым фактором, оказывающим влияние на растворимость белков, является их нативная конформация в растворе. Правильное сворачивание (фолдинг) полипептидных цепей некоторых белков в клетках эукариот обеспечивается специфическими белками, называемыми шаперонами, которые необходимы для эффективного формирования третичной структуры полипептидных цепей других белков, но не входят в состав конечной белковой структуры. Шапероны оказывают влияние лишь на конформацию полипептидных цепей и не действуют на ковалентные связи белковых молекул. К этому классу полипептидов относятся цитозольные белки теплового шока Hsp70 и Hsp60, а также гомологичные им белки, например белок, связывающийся с тяжелой цепью иммуноглобулинов (BiP). Шапероном является также ядерный нуклеоплазмин, обеспечивающий сборку нуклеосом. У E. coli соответствующие функции выполняют белки SecB, триггерный фактор, а также GroEL и GroES, обеспечивающие экспорт полипептидов из цитоплазмы и участвующие в морфогенезе бактериофагов. Два последних белка – GroEL и GroES, известные также как Cpn60 и Cpn10, кодируются генами оперона groE, регулируемого тепловым шоком. Белки, эволюционно родственные белку GroEL, получили название шаперонинов. Один из таких белков, обнаруженный в хлоропластах высших растений, участвует в сборке D-рибулозо-1,5-дифосфаткарбоксилазы-оксигеназы (Rubisco), которую часто используют в качестве модельного белка в исследованиях механизмов сворачивания полипептидных цепей.
Из других белков, участвующих в сворачивании полипептидных цепей, следует упомянуть так называемые фолдазы и изомеразы. Наиболее хорошо изученными ферментами этой группы являются тиоредоксин и глутаредоксин, которые наряду с глутатионзависимым превращением рибонуклеотидов в дезоксирибонуклеотиды способны восстанавливать дисульфидные связи в белках, способствуя формированию у них правильной третичной структуры. Те же функции выполняет и дисульфидизомераза белков, осуществляя образование дисульфидных связей в секретируемых белках после их переноса через мембраны эукариотических клеток. Лимитирующей стадией в образовании правильной третичной структуры полипептидных цепей является цис-транс-изомеризация остатков пролина в полипептидных цепях. Этот процесс в клетках эукариот осуществляется с участием пролил-цис-транс-изомеразы. Поскольку все вышеперечисленные условия, необходимые для поддержания эукариотических рекомбинантных белков в растворимой форме, не могут быть реализованы в бактериальных клетках, в них происходит агрегация рекомбинантных белков с образованием телец включения. В настоящее время не существует универсального метода, который бы позволял получать рекомбинантные эукариотические белки в бактериальных клетках в нативном виде. В некоторых случаях уже понижение температуры выращивания рекомбинантных бактерий до 30o приводит к образованию полностью растворимых рекомбинантных белков. Это наблюдали в случае рекомбинантного -интерферона, A-цепи рицина, щелочного фактора роста фибробластов и в ряде других случаев.
Более универсальным подходом к получению рекомбинантных белков в растворимой форме является обеспечение секретируемого характера их экспрессии, т.е. секреции синтезирующихся рекомбинантных белков в питательную среду. Системы секреции у грамположительных и грамотрицательных бактерий интенсивно изучаются. Получение рекомбинантных эукариотических белков в секретируемой форме высокотехнологично, поскольку значительно облегчает их очистку. Кроме того, у секретируемых белков больше шансов приобрести нативную конформацию, так как сворачивание полипептидных цепей происходит в большом объеме, а следовательно, и более низкой их концентрации, что предотвращает агрегацию полипептидных цепей, не полностью сформировавших свою третичную структуру. Примером эффективного использования секреторной системы E. coli является получение рекомбинантного инсулино-подобного фактора роста I (IGF-I) в секретируемой форме. В этой системе удавалось нарабатывать до 1 г рекомбинантного белка в 1 л бактериальной культуры.
Еще одним возможным, хотя и более труднореализуемым подходом к эффективному синтезу растворимых рекомбинантных белков в бактериальных клетках является введение экспрессирующихся генов шаперонов в клетки бактерий-хозяев. Так, сверхэкспрессия гомологичного gro-оперона в клетках E. coli, содержащих экспрессирующиеся гены большой и малой субъединиц Rubisco цианобактерии Anacistis nidulans, приводила к 5–10-кратному возрастанию внутриклеточного содержания нативного рекомбинантного белка Rubisco. В некоторых случаях внутриклеточную агрегацию сверхэкспрессирующихся белков удается предотвратить искусственной заменой отдельных аминокислот в полипептидных цепях методами белковой инженерии. Например, удаление семи аминокислот перед спиралью G в N-концевой части полипептидной цепи фрагмента Кленова ДНК-полимеразы I E. coli, а также замена нескольких гидрофобных аминокислот, боковые цепи которых экспонированы на поверхности спирали G, на остатки аспарагина полностью предотвращали внутриклеточную агрегацию фрагмента и образование агрегатов в процессе его очистки. В нативной молекуле ДНК-полимеразы I участок полипептидной цепи, в котором проведены замены, закрыт доменом 5’3’-экзонуклеазы, отсутствующим у фрагмента Кленова, и не оказывает влияния на агрегационные свойства фермента.
Несмотря на определенные успехи в получении нативных рекомбинантных эукариотических белков в бактериальных клетках, эти системы экспрессии эукариотических генов в настоящее время еще далеки от совершенства. По-прежнему остается открытым вопрос о посттрансляционных модификациях рекомбинантных белков в бактериях, а для многих рекомбинантных белков не решена даже проблема их внутриклеточной растворимости. С этой точки зрения идеальными системами экспрессии рекомбинантных эукариотических генов являются гомологичные генетические системы, т.е. сами эукариотические клетки животных или растений (см. раздел 7.4). Действительно, такие системы экспрессии, хотя и более дорогие, с успехом используются в биотехнологии. При этом одним из ключевых факторов, обеспечивающих эффективное функционирование подобных систем, могут быть эукариотические экспрессирующие векторы, основные особенности которых будут рассмотрены в следующем разделе.
7.2.7.Векторы для переноса ДНК в клетки животных и растений
Все основные принципы, используемые при конструировании бактериальных векторов, применимы и для получения векторов эукариотических клеток. Как и в случае бактерий, эукариотический вектор представляет собой небольшую молекулу ДНК, способную автономно реплицироваться в клетках животных. Помимо последовательностей нуклеотидов, обеспечивающих репликацию, эукариотические векторы могут содержать гены, используемые в качестве селектируемых маркеров, а также один или несколько уникальных сайтов рестрикции, по которым производится встраивание клонируемых последовательностей нуклеотидов ДНК. Поскольку непосредственное клонирование рекомбинантных ДНК в клетках животных было бы дорогостоящей и малоэффективной процедурой, эукариотические векторы используют, как правило, для получения экспрессии уже клонированных последовательностей нуклеотидов в клетках животных и растений, а сам процесс клонирования проводят в бактериях. Следовательно, эукариотические векторы, помимо всего прочего, должны быть челночными векторами. Для экспрессии в клетках рекомбинантные ДНК помещают под контроль регуляторных элементов, узнаваемых и используемых ферментативными системами эукариотических клеток.
В отличие от бактерий в клетках эукариот внехромосомные, автономно реплицирующиеся генетические элементы типа бактериальных плазмид встречаются редко. Поэтому при конструировании векторов, способных реплицироваться и осуществлять экспрессию в клетках животных, чаще всего используют регуляторные генетические элементы вирусов животных или, в зависимости от задач, эукариотических генов домашнего хозяйства, а также генов, для которых характерна тканеспецифическая экспрессия.
Экспрессирующий вектор pKSV-10. Структура теоретически возможного эукариотического вектора для экспрессии клонированных генов в клетках животных представлена на рис. II.12, из которого видно, что эукариотический вектор сохраняет все основные генетические элементы бактериальных векторов. Это, прежде всего, репликатор (область начала репликации ori), распознаваемый репликативными системами эукариотической клетки. В роли репликатора чаще всего используют соответствующие последовательности нуклеотидов вирусов животных (например вируса SV40 или вируса Эпштейна–Барр). Особенностью функционирования такого репликатора является потребность в вирусных белках для инициации репликации. Так, в случае SV40 – это T-антиген, тогда как у вируса Эпштейна-Барр – белок EBNA (Epstein-Barr nuclear antigen). Для того чтобы репликация векторов, в которых использованы вирусные репликаторы, осуществлялась без вируса-помощника, получили специальные линии клеток, которые стабильно продуцируют соответствующие вирусные белки. Например, в клетках линии COS синтезируется T-антиген вируса SV40, а клетки линии Hep-EBNA-2 экспрессируют ген EBNA.
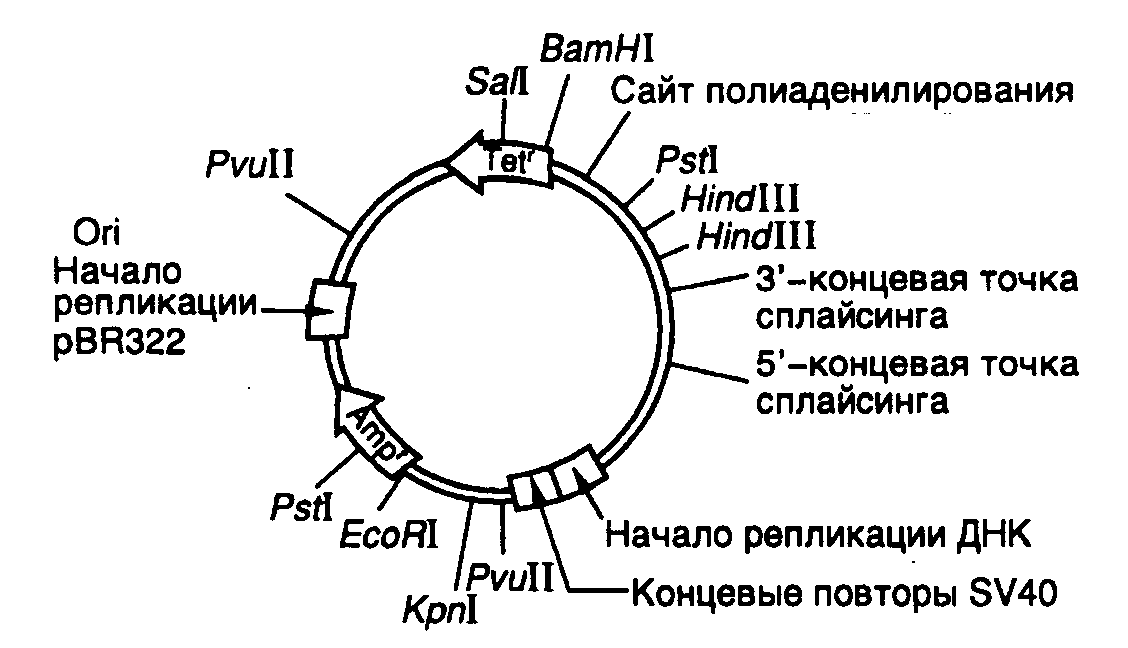
Рис. II.12. Эукариотический вектор pKSV-10
Часть бактериальной плазмиды, содержащей область начала репликации ori, соединена с транскрипционной единицей вируса SV40, 5’-концевая часть которой включает ранние промотор и репликатор вируса SV40, а 3’-конец содержит сайт полиаденилирования и интрон большого T-антигена. Последовательности чужеродной ДНК, которые необходимо экспрессировать в клетках животных, клонируют по BglII-сайту рестрикции. Кроме того, в векторе имеются уникальные сайты для рестриктаз KpnI, BamHI, SalI и EcoRI. Вектор может реплицироваться в клетках E. coli и в клетках обезьян линии COS
В ряде случаев гены, необходимые для репликации эукариотического вектора, вводят непосредственно в вектор. Так же как и бактериальный, эукариотический вектор должен содержать один или несколько уникальных сайтов рестрикции для клонирования требуемой последовательности нуклеотидов. В экспрессирующем векторе полилинкер располагают непосредственно за генетическими элементами, регулирующими транскрипцию (наибольшее значение из них имеют промотор и энхансер), перед сайтом полиаденилирования и терминатором транскрипции. Структура других кассет генов, обеспечивающих экспрессию рекомбинантных генов в клетках животных, будет рассмотрена в разделе 7.4.6.
Векторы растений. Специфика векторов для экспрессии рекомбинантных генов в клетках растений обусловлена главным образом регуляторными последовательностями нуклеотидов, которые обеспечивают эффективную транскрипцию генов в растительных тканях. Например, в известном векторе pCaMVCAT используются промотор и сайт полиаденилирования генов вируса мозаики цветной капусты, под контролем которых находится ген бактериальной хлорамфениколацетилтрансферазы (cat). Этот ген-репортер применяется для оптимизации условий введения векторной ДНК в клетки растений и может быть заменен на любой другой ген. Вектор pCaMVCAT является челночным, т.е. может реплицироваться как в клетках E. coli, так и растений. В первом случае селектируемым маркером служит, как обычно, ген устойчивости к ампициллину.
Вектор pCaMVCAT принадлежит к большой группе так называемых DMGT-векторов (DNA-mediated gene transfer), осуществляющих опосредованный ДНК перенос генов в клетки растений. Это означает, что в данном случае перенос генов в клетки растений опосредован ДНК вектора, так как они находятся в составе векторной молекулы. Для получения стабильных линий растительных клеток, которые бы содержали рекомбинантные ДНК в интегрированном в хромосомы состоянии и активно экспрессировали его, часто в состав DMGT-векторов вводят дополнительные селектируемые маркеры, которые придают клеткам, включившим экзогенную ДНК, определенный, легко обнаруживаемый фенотип. Такими маркерами могут быть, например гены неомицинфосфотрансферазы II (NPT II), гигромицинфосфотрансферазы (HPT), дигидрофолатредуктазы, в результате экспрессии которых клетки приобретают устойчивость к канамицину, гигромицину B или метотрексату соответственно.
|
|
|
 Скачать 5.83 Mb.
Скачать 5.83 Mb.