Цитология методичка 1-15 сабак. Саба 1 Апаратты дидактикалы топтама
 Скачать 1.99 Mb. Скачать 1.99 Mb.
|

Репликацияның негізгі ферменттері:ДНҚ-полимераза – ДНҚ тізбегін синтездейді топоизомераза – репликациялық айыр алдындағы ДНҚ-ың суперспирализациясын босаңсытады, тарқатады хеликаза -ДНҚ тізбектерін ажыратады SSB ақуыздары – ажыраған бір тізбекті ДНҚ бөліктерін тұрақтандырады лигаза – ДНҚ фрагменттерін тізбектеп тігеді РНҚ-праймаза - ДНҚ- полимераза үшін РНҚ-бастаушты (праймерлер) синтездейді 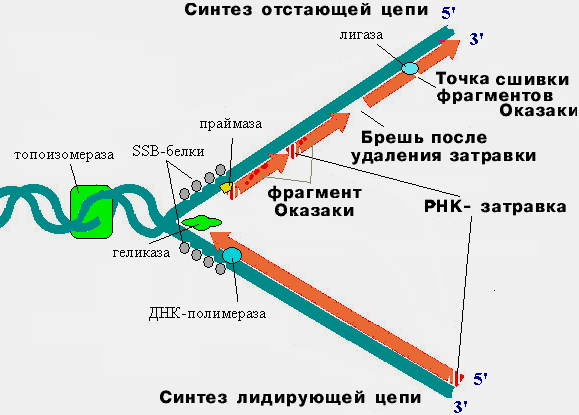 Репликациялық айыр аймағындағы ДНҚ репликацииясының сызбасы Репликацияның өте жоғары дәлділігі қосымша ДНК-полимеразамен қамтамасыз етіледі. Бұл ферменттің қызметі ДНҚ комплементарлық ұстанымдарын және дұрыс емес нуклеотидтерді алып тастайды түзету және жөндеу жүргізеді. Прокариоттарда секундына 3000 жұпқа жуық нуклеотидтер, ал эукариоттарда - секундына 100-300 жұп нуклеотидтер синтезделеді. Дегенмен де репликация кезіндегі гендік (нүктелік) мутациялар – азотты негіздердің алмасуы, репликация қателіктеріне әкеледі. Уотсон және Крик қос спиральды ДНҚ моделінің негізінде ДНҚ-ң азоттық негіздерінің алмасуының алғашқы механизмін ұсынған. Олардың көз қарастары бойынша нуклеотидтер арасындаға комплементарлы A-T және G-C жұптары арасында таутомеризаця жүруі де мүмкін, бұл жағдайда комплементарлық конформациясы өзгереді, аденин гуаниннің қызметіне, гуанин-аденинге, цитозин-тиминге, тимин-цитозинге ауысады. Сонымен қатар репликацияның бұзылу себептеріде әртүрлі болуы мүмкін. Мысалы, ультракүлгін сәуленің әсерінен нуклеотидтер концентрациясының өте жоғары немесе өте төмен болуы, пуриндік негіздердің спонтанды түсіп қалуы, цитозиннің дезаминденуі себебінен урацилге айнала алады. Пайда болған қателіктер мен бұзылыстар репарациялық үдеріспен және ДНҚ-полимеразаның жөндеу және түзету жүргізе алу қабілетіне байланысты қателіктер түзетіліп отырады, ал кейбір жағдайларда ДНҚ молекуласындаға қателіктер түзетілмей қалып қояды. Егер зақымданулар ДНҚ-ң функциональды активті емес аймағында өтетін болса, онда ондай қателіктер фенотиптік көрініс бермейді және ешқандай зілді қауіптер байқалмайды, бірақ қандайда бір генде нуклеотидтер қатарының жұптасуында қателіктер кететін болса, онда фенотиптік өзгерістің көрініс беру мүмкіндігі көбее түседі де гендік аурулардың пайда болуына әкеледі. Репликация үдерісіне әртүрлі сыртқы орта факторлары, соның ішінде дәрілік заттарда әсер етеді. Мысалы, дауномицин және ісікке қарсы препараттар ДНҚ құрылысын өзгертеді, нәтижесінде репликация ферменттері жұмыстарын тоқтатады. Мысалы, топоизомераза ДНҚ босаңсыту (деспиралдандыру) мүмкіндігінен айырылады. Алкилдеуші заттар тиофосфамидтер де ДНҚ молекуласындағы азотты негіздердің модификациясын тудырады да репликация ферменттерінің жұмысын тоқтатады. ДНК- топоизомеразасының ингибиторы-новобиоцин ферменттер жұмысына араласып ДНҚ деспиральдануын тоқтатады. Сонымен репликация үдерісі генетикалық ақпараттың берілуінің негізгі кезеңі, ондағы бұзылыстар жасушаның ары қарай бөлінуін тоқтатуы мүмкін. Сол сияқты репликация жылдамдығының (төмендеуі) өзгеруі мүшелердің дамымай қалуына, эмбриогенезде репликацияның жоғарлауы мүшенің гиперплазиясына және гендік аурулардың пайда болуына себепші болады. ДНҚ молекуласындаға генетикалық ақпарат рибонуклеин қышқылының (РНК) молекуласына беріледі. РНҚ-ң ДНҚ-дан айырмашылығы қант молекуласы дезоксирибозаның орнында рибоза және азотты негізі - урацил (тиминнің орнына) аденинге комплментарлы. ДНҚ транскрипциясы инициация, элонгация және терминации сатыларынан тұратын ДНҚ екі тізбегінің бірінен РНҚ молекуласы синтезделетін матрицалық үдеріс. Инициация - алғашқы нуклеотид аралық байланыс РНҚ-полимеразаның промоторымен байланысуынан басталады. Осы кезде белгібір жерден ДНҚ тізбегі тарқатылып транскрипциялық «көзше» пайда болады. Прокариоттарда РНҚ-полимераза Прибнов бокстың нуклеотидтер қатарындағы s(сигма) – факторды таниды, соңынан оған екі a тізбегінен, b(бета) b¢(бета штрих) және w (омега) суббірлігінен тұратын кор-фермент қосылады. Бұл кешен холофермент деп аталып транскрипцияның басталу нүктесіне қарай жылжиды, бірнеше нуклеотидтер транскрипцияланғанан кейін, s фактор ферментімен байланысын үзеді, тек қана кор-фермент ДНҚ бойымен жылжиды. Эукариоттардың РНҚ-полимеразасы өздігінен транскрипцияны инициациялай алмайды, оған жалпы транскрипциялық фактор-ақуызы (ЖТФ) қажет. Ол РНК-полимеразаны ТАТА-бокспен байланыстырады және бұл кешен (РНҚП+ЖТФ) транскрипцияның басталу нүктесіне қарай жылжиды. Элонгация - РНҚ тізбегінің ұзаруы, транскрипцияның басталу нүктесінен басталып және терминациялық сайтта униполярлы, консервативті және комплементарлы ұстаныммен аяқталатын үдеріс. Терминация- транскрипцияның аяқталуы, геннің терминациялық сайт қатарындағы ГЦ- ға бай бөлігінде бір тізбекті РНҚ молекуласының бөлініп шығуымен аяқталады. ДНҚ әрбір бөлігінде транскрипцияның аяқталуынан кейін екі тізбекті ДНҚ молекуласы бастапқы қалпына келеді. Транскрипция үдерісіне арнайы фермент РНҚ-полимеразамен қатар көптеген инициациялық, элонгациялық және терминациялық реттеуші ақуыз факторлары да қатысады. Мұндай ақуыздар транскрипцияның басталу және аяқталу үдерісін және алғашқы өнімнің деңгейінің реттелуін қамтамасыз ететін гендердің реттеуші қатарларымен байланыс түзеді. Эукариоттар транскрипциясының ерекшелігі де сол активті емес ядролық РНҚ түзіледі және постранскрипциялық модификация-процессингті (пісіп-жетілу) жүргізу болып табылады. Процессинг үдерісінде жүреді: «кэп»- 51ұшына–метилденген гуанозиннің қосылуы полиадениннің- 31 полиадениндік «құйрықтың»жалғануы сплайсинг- интрондардың қырқылуы және экзондардың тігілуі Посттранскрипциялықмодификация Транскрипцияның алғашқа өнімі немесе ядролық РНҚ, цитоплазмалық РНҚ қарағанда активті емес (жетілмеген) және одан әлде қайда ұзын болады. Жетілмеген ядролық РНҚ құрамында ақпараты бар, кодталатын қатарлардан (экзондардан) және ішінде ақпараты жоқ немесе кодталмайтын қатардан (интрондар) тұрады. Транскрипцияның алғашқа өнімі а-РНҚ, гетерогенді ядролық РНҚ (гя РНҚ) деп аталады немесе про- а-РНҚ, онда 500 - 50000 нуклеотидтер қатары болады. Гя-РНҚ әртүрлі өзгерістерге ұшырауы мүмкін: Кейбір гендердің транскрипты РНҚ-ы цитоплазмаға өтпейді, жетілген цитоплазмалық РНҚ айналмайды, ядрода-ақ жойылады. Алғашқы әрбір геннің РНҚ транскрипты толықтай процессингке ұшырамайды, тек ядролық РНҚ біраз ғана (25%) бөлігі жетілгена-РНҚ айналады. Қалған 75% ядрода жойылады. Сонымен тек барлық РНҚ тек 5% ғана цитоплазмаға өтеді. Ядролық РНҚ жетілген функциональды активтіа-РНҚ айналуы және цитоплазмаға немесе процессингте іске асады: 1. Ядролық РНҚ-ның цитоплазмаға өтіп, жетілген, функциональды активті а-РНҚ-ға айналуы немесе процессинг мына әрекеттерді қамти жүреді: «Қалпақ» жалғану немесе «кэп» жалғану. «Кэп» жалғану үдерісі пре-а-РНҚ-ның 51 – ұшына метилденген гуанозиннің жалғануы. Мұны 7 метилгуанин (7-МеГ немесе m7Г) деп атайды, ол 5/ – 5/ арасындаға сирек байланысты түзеді. «Кэп» жалғану РНК – полимераза II транскрипцияның инициация сайтынан ажырауына дейін және нуклеотидтердің қырқылуы мен сплайсинг үдерісінің басталуына дейін басталады. Кейде «Кэп» жалғану нуклеотидтердің қант молекуласының қосымша метилденуі 5/-ұшына (7-Мег-5/-РРР-5/), рибозаның 3/ ұшының Г немесе А метилденуі моно- және трифосфаттануына сай келеді. «Қалпақ» жалғанудың биологиялық маңызы «сар»-қалпақ а-РНҚ –ны нуклеаза ферменттерінің бұзу әрекетінен қорғайды, сонымен қатар ақуыздың биосинтезіне қатынасатын факторлардың а-РНҚ-ны танып байланысуына көмектеседі. Мұндай 7-МеГ- «қалпақ» гистонды белоктардың а-РНҚ-да болмайды. Полиадениндік «құйрықтың» жалғануы, а-РНҚ-ның 3/ұшына поли-А-фрагменттің жалғауы екі кезеңнен тұрады: а) эндонуклеаза ферменті пре-а-РНҚ-ны (бірінші реттік транскриптті) белгілі учаскеде кеседі. б) пре-а-РНҚ-ның кесілгеннен кейінгі түзілген 31 ұшына полиаденилдік фрагмент («құйрық») жалғанады. Бұл үшін арнайы фермент –поли-(А)-полимераза қатынасып субстрат ретінде АТФ-ты пайдаланады, процестің өзін полиаденилдену деп атайды. Жалғанған полиаденилдік фрагмент шамамен 200-ге жуық нуклеотидтен тұрады. Әдетте полиаденилдену пре-а-РНҚ-ның 30% қамти жүрсе, жетілген а-РНҚ тізбегінің -70% жуығын қамтиды. Көптеген а-РНҚ гистонды ақуыздары 31ұшында полиаденилдену жүрмейді яғни процессинг поли – А жалғанусыз өтеді. Процессинг кіші ядролық РНҚ У (У7 snRNA) 57 жұп негіздер экстенсивті комплементарлы жұптасуы гяРНҚ гистонды ақуыздарымен байланысы арқалы іске асырылады. РНҚ сплайсингі - алғашқы транскриптан кодталмайтын қатардың немесе интронның қырқылуын және РНҚ-ың қалған фрагменттерінің, кодтаушы қатардың немесе экзондардың тігілуінен жетілген функциональдық активті а-РНҚ молекуласының түзілуін бақылайтын үдеріс. Пре-а- РНҚ-дағы интрондардың қырқылып түсуі мен экзондардың жалғану механизмдері жайлы нақты дәйекті мәліметтер әлі де жеткіліксіз. Әр интронды учаскенің басы мен аяғында нақты консенсустық нуклеотидтер жүйесі орналасқан: кез-келген интрон әрдайым Г-У-мен басталып, А-Г дуплетімен аяқталады. Екіншіден, бұл нуклеотидтік жүйелерді дұрыс танып табу үшін арнайы кіші ядролық РНҚ-лардың белоктық бөлшектері (sn RNP) қатысады. Сплайсеосоманың сол жақ (ГУ) ұштарын кіші ядролық РНҚ-ң У1, ал оң жақ ұшын кіші ядролық РНҚ-ң У5 таныйды. Кіші ядролық РНҚ интрондар ішінде болатын басқада консенсустық «жеке сайтты», қатарды таниды. Кіші ядролық РНҚ-ң У4 және У6 сплайсеосомалар түзуге қатысады, бірақ олардың ролі әлі белгісіз. Сплайсинг механизмі аса дәлдікпен жүруі тиіс, интрондың қырқылуы мен экзондардың тігілуінде кететін қателіктер тігілген кодондардың кодталуының бұзылуына әкеледі, соның нәтижесінде мутантты ақуыздың түзілуі немесе оның сиинтезделмеуі мүмкін. Сплайсингтің молекулалық механизмі жетілген РНҚ молекуласына бастама беретін РНҚ (про- а-РНҚ, гетерогенді ядролық РНҚ) бірінен соң бірі ретпен жүретін күрделі үдерісі. Кіші ядролық РНҚ (мяРНҚ) және ақуыздар сплайсинг үдерісіне әсер ете отырып тиімділігіне әсер етеді. Кіші молекулалық РНҚ-ң гя-РНҚ молекуласымен бірігуі (про –а- РНҚ) Г – Ц және А – У комплементарлы ұстанымы бойынша іске асады. Интрондардың қырқылып алынып тасталуы жүреді, егер экзондар шекарасында бір жағында алмаспайтын ГУ (консенсусты) нуклеотидтері екінші жағында АГ болатын болса. Нүктелік мутация нәтижесінде бұл нуклеотидтердің алмасуы жүретін болса (мысалы гуаниннің (Г) адининге (А) сплайсинг бұзылады, мутантты ақуыз түзіледі немесе ақуыз түзілмейді. Мысалы, мұндай мутация адам гемоглобинінің генен орын алған, тұқым қуалайтын анемияға әкеледі. Экзондар шекарасындағы нуклеотидтер арасындағы үзілістер экзон 1 мен интрон арасында аденинмен интронның бірінші нуклеотидінің (Г) байланысы ілмектің пайда болуына әкеледі, нәтижесінде интрондар алынып экзондар бір-бірімен тігіледі. Экзондардың сплайсингінің арасында кіші ядролық (мя РНК) РНҚ үш типті 2,5, және 6 болады. Кіші ядролық (мя РНК) РНҚ молекуласында жұптаспаған нуклеотидтер 5 РНҚ қос спиральды ілмегін түзеді. Ілмек нуклеотидтері экзондармен сутекті байланыс түзіп, бірін-бірі жақын ұстап тұрады. Кіші ядролық РНК 2 (мя РНК) молекуласы интрондар арасындағы байланысты ұстай отырып, мяРНҚ 6 бөлігімен «жұптасады». Аралық құрылымның ерекшелігі де сол, интрондар шетіндегі екі гуаниндер арасындағы байланысты түзеді. Мұндай байланыстар экзондарды ұстап тұрады да сплиайсингті қамтамасыз етеді. Кіші ядролық РНК 6 (мя РНК) сплайсингтің белгілі бір кезеңінде активті болады. Сплайсинг үдерісіне ферменттік қызмет атқаратын бірқатар белоктар қатынасады: энергия көзі ретінде қызмет атқаратын белоктар АТФ-азалар; РНҚ-геликазалар – сплайсинг барысында түзілген қосарлана ширатылған РНҚ-РНҚ тізбектерді тарқатуды және кіші ядролық РНҚ-лар арасындағы РНҚ-РНҚ-лық (мысалы, мя РНК 4- мя РНК 6) өзара әрекеттесулерді бұзу арқылы ДНҚ молекулаларының арасында жаңа байланыстардың түзілуін қамтамасыз ететін ферменттер. Олай болса, сплайсинг күрделі үдеріс, оның реакциялары рибонуклеотидтік ірі кешенде, бірнеше РНҚ молекулалары және ондаған белоктардың қатынасуымен жүреді. |