Навигация по странице:/ — мочевой пузырь, 2 — выделительная пора, 3, 4, 5 — различные участки канала, 6 — целомический мешочек/ — энтодерма, 2— первичные мезодермальные клетки/ — антенна, 2 —мандибула, 3— ан- теннула, 4 — науплиальный глаз. 5 — антенпальная выделительная железа, 6 — кишечник 1 — антеннула, 2 — антенна, 3 — мандибула, 4 — науплиальный глаз 1 — сложный глаз, 2— антеннула, 3 — антенна, 4, 5 — ногочелюсти, 5 — зачатки грудных ног, 7 — брюшко, 8 — последняя пара брюшных ножек/ — антеннула, 2— антенна, 3 — грудные конечности, 4 — брюшные ножки, 5 — хвостовая плавательная ножка, 6 — тельсон
|
В.А. Догель - Зоология беспозвоночных. Тип губки (spongia, или porifera)
Branchipus (из Клауса); В — участок разреза через сложный глаз: / — ретинальные клетки, 2 — зрительный нерв, 3—хрусталик, 4 — пигментные клетки, 5 — хрустальные конусы, 6 — слой ретинальных и пигментных клеток, 7 — нервные волокна, 8 — мускул глазного стебелька, 9 — мозг. 10, II — ганглиоз-ные скопления нервных клеток, 12 — хитиновая прозрачная кутикула, 13 — ди-стальные части ретинальных клеток, 14 — ретинальные клетки, 15 — базальная мембрана, 16 — нервные волокна, отходящие от базальной части ретинальных клеток, 17 — прослойки пигмента между отдельными омматидиями
сти тела, занимает светопреломляющий хрусталик. Глаз имеет инвертированное строение, так как нервные волокна отходят от концов ретинальных клеток, обращенных в полость глазного бокала.
Во-вторых, большинство раков обладает парой сложных, или фасе-тированных, глаз (рис. 266,5), очень характерных для большинства членистоногих. Каждый такой глаз содержит множество мелких глазков, или омматидиев (у речного рака их больше 3000), тесно сближенных и отделенных друг от друга лишь тонкими прослойками пигмента.
304
Омматидий (рис. 266.В), в свою очередь, состоит из сложно устроенного светопреломляющего аппарата, группы чувствительных ретинальных клеток, от которых отходят нервные окончания, дающие начало зрительному нерву, и экранирующих пигментных клеток'.
Сложные глаза часто сидят на особых подвижных выростах головы, стебельках. У различных представителей раков могут быть во взрослом состоянии следующие комбинации глаз: 1) только науплиальный глаз
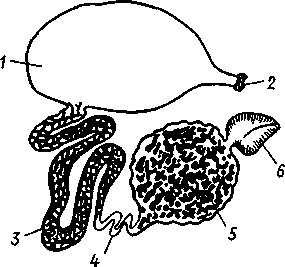  * *
Рис. 267. Действие гормона синусовой железы на хроматофоры креветки Le-ander. A — пигментные клетки; Б — посветлевшие под влиянием гормонов пигментные клетки (из Ганшт-рема)
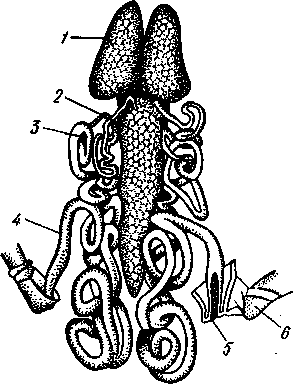
Рис. 268. Антеннальная железа речного рака (из Вальса):
/ — мочевой пузырь, 2 — выделительная
пора, 3, 4, 5 — различные участки канала,
6 — целомический мешочек
(отр. Copepoda, большинство представителей подкл. Ostracoda), 2) только сложные глаза (многие высшие раки), 3) и те и другие вместе (многие листоногие раки и др.). Интересно, что у некоторых ракообразных зрительные восприятия определенных световых раздражений необходимы для побуждения ней-росекреторных клеток к выделению гормонов, влияющих на окраску тела. Давно известна способность некоторых креветок и крабов временно изменять цвет тела в зависимости от фона окружающей среды. Способность эта зависит от перемещения зерен окрашенного пигмента в цитоплазме особых крупных клеток-хроматофоров, находящихся под покровами. Когда зерна равномерно рассеяны по всей клетке, она окрашена в тот или иной цвет в зависимости от цвета пигмента, при концентрации последнего в центре клетки она бледнеет (рис. 267). Деятельность пигментных клеток регулируется нейросекретом, продуцируемым синусовыми железами (см. рис. 263). Таким образом, зрительные раздражения через нервную систему действуют на работу синусовых желез, а выделяемые ими гормоны распространяются кровью по всему телу и вызывают (на светлом грунте) концентрацию пигментных зерен в хромато-форах, приводящую к общему посветлению тела. Такое изменение окраски — приспособление защитного характера. Выделительная система раков почти утратила метамерный характер. Имеются 2 пары выделительных железистых органов — видоизмененных целомодуктов. Строение обеих пар приблизительно одинаково. Каждый орган состоит из концевого мешочка и отходящего от него извитого канала с железистыми стенками; канал делает несколько петлеобразных 1 Более подробно вопрос о строении сложного глаза и его функциональных особенностях будет рассмотрен в главе о насекомых — с. 360. 305
изгибов и затем открывается наружу, иногда образуя перед этим заметное расширение — мочевой пузырек (рис. 268). Одна пара открывается у основания антенн (антеннальные железы), другая — у основания второй пары нижних челюстей (максиллярные железы). Установлено, что концевой слепо замкнутый мешочек представляет собой сохранившийся от эмбрионального состояния участок целома, а отверстие, сообщающее пузырек с каналом,— воронку целомодукта, только лишенную мерцательных ресничек. Лишь в очень редких случаях (отр. Leptostraca) обе пары выделительных желез присутствуют одновременно. Обыкновенно же одна из них функционирует на личиночных стадиях и потом редуцируется, заменяясь во взрослом состоянии другой. При этом все раки, кроме подкл. Malacostraca, во взрослом состоянии имеют максиллярные железы, a Malacostraca — антеннальные железы.
Половая система. Громадное большинство раков раздельнополы. Нередко имеется явственный половой диморфизм. Так, у самцов либо ан-теннулы, либо антенны могут превращаться в хватательные органы для удерживания самки (см. рис. 277,5). Конечности, расположенные близко к половым отверстиям, становятся копулятивными органами: таковы I и II пары брюшных ножек речного рака (у самки I пара рудиментарна) (см. рис. 252). У низших форм самцы нередко значительно мельче самок. Иногда разница столь велика, что можно говорить о карликовых самцах, например, у некоторых паразитических форм, а также у ряда представителей отряда усоногих раков Cirripedia, ведущих сидячий образ жизни. Кроме того, у последних часто развивается настоящий гермафродитизм (с. 319).
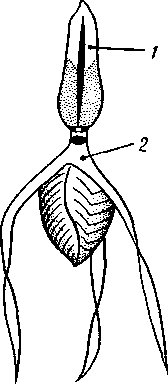
В наиболее примитивных случаях половые железы парны: это два мешка, сообщающиеся с внешней средой при помощи половых протоков. Гораздо чаще обе гонады полностью или частично сливаются, но протоки их всегда остаются парными (рис. 269). Яйцеводы имеют вид коротких трубок с железистыми стенками, выделяющими вокруг яиц плотную скорлупу. Нередко у самок имеются еще особые семяприемники, лежащие вблизи, но независимо от женских половых отверстий. Через специальное копулятивное отверстие в семяприемники вводится при половом акте семя самца и хранится там до момента выхода яиц из женских половых отверстий, когда, собственно, и наступает оплодотворение. Семяпроводы иногда расширяются в семенные пузырьки и тоже обладают железистыми стенками. Выделениями последних живчики склеиваются в большие, одетые оболочкой пакеты — сперматофоры. При копуляции сперматофоры или вводятся самцом в половые отверстия самки, или только подвешиваются к ним. Положение половых отверстий у раков варьирует: у всех высших раков они находятся у самок на 6-м, а у самцов на 8-м грудном сегменте (см. рис. 252).
Большинство раков обнаруживает заботу о потомстве, вынашивая яйца приклеенными либо к половым отверстиям (яйцевые мешки отр. Copepoda; см. рис. 282),либо к конечностям брюшка (отр. Decapoda) и т. п. Плодовитость раков различна.
У раков, обладающих типичными живчиками, оплодотворение совершается обычным путем. Отметим колоссальную длину сперматозоидов у некоторых раков; у представителей отр. Ostracoda длина живчиков достигает 6 мм (самые длинные сперматозоиды во всем животном мире), превышая в 10 раз длину самого животного.
У многих Malacostraca, однако, сперматозоиды имеют причудливую форму и совершенно неподвижны (рис. 270). Так, у Galathea (из отр. Decapoda) живчик состоит из двух овальных мешков (один из них на конце заострен), соединенных узким пояском, от которых отходят в одну
306
сторону 3 длинных отростка. Головке обыкновенных спермиев отвечает вздутая и заостренная половина, хвостовому отделу — другой овальный мешочек, или «хвостовая капсула». Попадая в половые протоки самки, живчики зацепляются за проходящие мимо яйца своими отростками и устанавливаются на поверхности яйца, как на треножнике, направляясь
Рис. 269. Мужской половой аппарат речного рака (из Гексли):
/ — парная часть семенника, 2 — непарная часть семенника, 3 — семяпровод, 4 — се-мявыносящий канал, 5 — половое отверстие, 6 — основание задней грудной ноги
Рис. 270. Живчик десятиногого рака Galathea. Головка живчика обращена книзу (по Кольцову):
1 — хвостовая капсула, отвечающая хвостовой нити обыкновенных живчиков, 2 — шейка с тремя отростками
головкой в сторону яйца. В это время на заднем конце хвостовой капсулы открывается особое отверстие, через которое окружающая жидкость проникает внутрь капсулы. Внутри нее находится студенистое вещество, которое жадно впитывает влагу, разбухает и в конце концов с силой выстреливается из заднего отверстия, не помещаясь более внутри капсулы. Это важный момент: вследствие происшедшего «взрыва» капсулы живчик получает обратный толчок и вонзается заостренной головкой в яйцо. После этого хвостовая капсула и отростки отбрасываются, головка же целиком проникает внутрь яйца.
Развитие. Яйца ракообразных обычно богаты желтком (рис. 271). Когда желтка мало (некоторые Copepoda и др.), дробление бывает полное, неравномерное и детерминированное, чем напоминает дробление кольчатых червей. В таких случаях еще на очень ранних стадиях одна из клеток, делясь, дифференцируется на клетку, дающую начало энтодерме, и на мезодермальный телобласт.
Однако у большинства раков обилие желтка изменяет характер дробления; оно становится частичным и поверхностным (рис. 271). Ядро яйца последовательно делится на 2, 4, 8 и более ядер без соответствующего деления самой клетки. Эти ядра уходят затем на периферию яйца, располагаются там в один слой, и вокруг каждого ядра обособляется участок цитоплазмы в виде небольшой клетки. Центральная масса
307
желтка остается неразделенной, и только поверхность ее одета одним слоем клеток. Отсюда частичное дробление такого типа называется поверхностным. Такая стадия отвечает бластуле, первичная полость которой заполнена желтком. Часть клеток бластулы на будущей брюшной стороне зародыша уходит под наружный слой и дает начало энтодерме
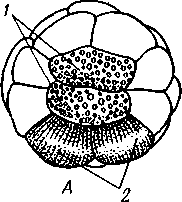 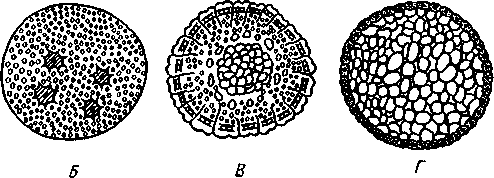
Рис. 271. Дробление яйца ракообразных (из Давыдова). А — детерминированное
дробление Euphausia; Б—Г — три стадии поверхностного дробления краба Dromia
(Г — стадия, отвечающая бластуле, но заполненная желтком):
/ — энтодерма, 2—первичные мезодермальные клетки
и мезодерме. Вследствие этого на брюшной стороне получается многослойная клеточная пластинка — зародышевая полоска. Поверхностный слой ее образован эктодермой, более глубокие слои представляют мезодерму, а самый глубокий слой, прилегающий к желтку, — энтодерму. Формирование зародыша происходит главным образом за счет зародышевой __ Л. полоски, начинающей сегментироваться, причем самый передний и мощный учас ток ее дает начало парным головным до лям, за счет которых возникают сложные глаза. Позади долей первыми заклады- 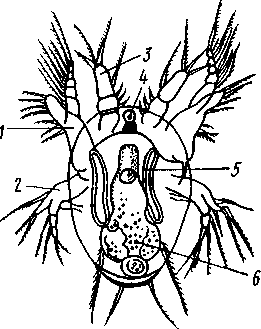
Рис. 272. Личинка науплиус
циклопа Cyclops (отр. Сореро-
da) (по Клаусу):
/ — антенна, 2 —мандибула, 3—ан-
теннула, 4 — науплиальный глаз.
5 — антенпальная выделительная
железа, 6 — кишечник
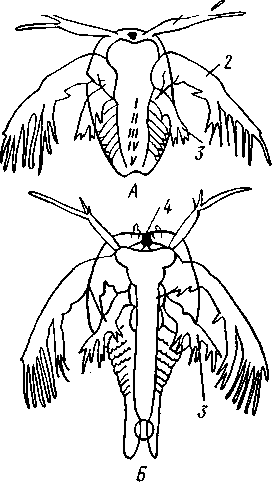
Рис. 273. Личинка щитня Apus. А — науплиус с зачатками пяти сегментов (I—V); Б — личинка с семью еще безногими сегментами (по Клаусу):
1 — антеннула, 2 — антенна, 3 — мандибула, 4 — науплиальный глаз
308
1
ваются зачатки головной лопасти (акрона) и двух передних сегментов1: антеннального и мандибулярного. Мезодерма зародыша в некотор^лХл случаях складывается в два ряда целомических мешков (как у кольча-тых червей), которые впоследствии, однако, разрушаются. Клетки их идут на построение мезодермальных органов (мышц, сердца и т. д.), а полости сливаются с остатками первичной полости тела, образуя полость смешанного происхождения — миксоцель. Иногда сегментация мезодермы у раков утрачивает свою отчетливость, а формирования ясного целома вообще не происходит.
Дальнейшее развитие у большинства ракообразных сопровождается метаморфозом различной сложности. У многих низших форм, например у листоногих раков — Phyllopoda, дело идет так. Зародыш развивает часть конечностей и выходит из яйцевой оболочки в виде личинки. Типичная исходная стадия метаморфоза — планктонная личинка науп-лиус (рис. 272), столь же характерная для раков, как трохофора для Polychaeta. Овальное тело науплиуса обнаруживает свою метамерию присутствием трех пар придатков: антеннул, антенн и жвал. Принадлежащие головной лопасти антеннулы с самого начала простые, тогда как прочие две пары, представляющие конечности, снабжены двумя ветвями. Антенны у науплиуса лежат еще на брюшной стороне тела, по бокам рта, и своими основаниями участвуют в собирании пищи; на переднюю сторону головы они перемещаются позднее, чем доказывается их происхождение от обыкновенных ходных конечностей, постепенно передвинувшихся вперед и изменивших также и функцию. Жвалы лежат позади антенн и сначала служат почти исключительно для плавания. Тело личинки, следовательно, слагается из головной лопасти (акрона), несущей глаз и антеннулы, и двух сегментов: антеннального и мандибулярного. Кроме того, сзади лежит еще анальная лопасть с порошицей.
Личинка имеет кишечник, головной мозг и два брюшных ганглия, науплиальный глаз и одну пару выделительных органов (чаще — антен-нальные железы). Между сегментом мандибул и анальной лопастью находится также несегментированная зона роста. За счет этой зоны идет образование недостающих сегментов тела. Науплиус растет в длину.
В зоне роста в направлении спереди назад намечаются границы между но-
7 /жшшг—7Z==^ вообразующимися сегментами (рис.
L^ /.ШШши^, <—^^ 273), а затем закладываются в виде
бугорков и их конечности. Так, сначала обособляются сегменты обеих пар
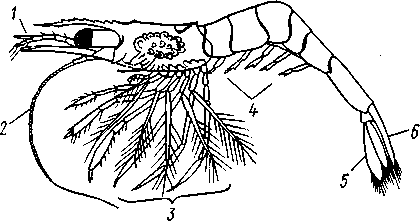
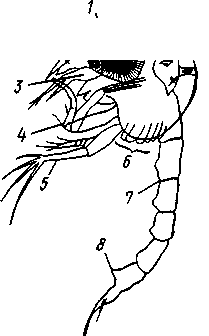
Рис. 274. Личинка зоеа краба Maja (по Клаусу):
1 — сложный глаз, 2—антеннула, 3 — антенна, 4, 5 — ногочелюсти, 5 — зачатки грудных ног, 7 — брюшко, 8 — последняя пара брюшных ножек
Рис. 275. Мизидная личинка креветки Рап-
dalus (по Сарсу):
/ — антеннула, 2—антенна, 3 — грудные конечности, 4 — брюшные ножки, 5 — хвостовая плавательная ножка, 6 — тельсон
309
нижних челюстей и передние грудные (стадия метанауплиуса), затем остальные грудные и позднее всего брюшные. Через известные промежутки времени личинка испытывает линьку, причем каждая линька приближает ее к состоянию взрослого животного. Постепенно дифференцируются парные глаза, конечности принимают окончательный вид, число сегментов становится полным, и метаморфоз заканчивается.
У высших раков в случаях наиболее примитивного и полного развития, например у некоторых креветок (из отр. Decapoda), процесс начинается так же. Из яйца выходит науплиус и постепенно превращается в метанауплиуса. Далее следует стадия, очень характерная для развития высших раков. Это зоеа (zoea). Так называют личинку (рис. 274) с развитыми ротовыми конечностями и ногочелюстями, а также с зачатками грудных ножек, сформированным брюшком, но имеющим пока лишь последнюю пару конечностей. Кроме того, стадия зоеа определяется по закладке пары фасетированных глаз. Следующая стадия метаморфоза характеризуется полным развитием грудных ног, которые имеют дву-ветвистую форму, и закладкой брюшных конечностей — мизидная стадия (рис. 275). После линьки мизидная личинка превращается в молодого сформированного рака.
Однако у значительного большинства представителей отр. Decapoda метаморфоз сильно укорачивается — часть его протекает внутри яйца. Так, у многих форм (например, у крабов) из яйца вылупляется прямо зоеа, а у речного рака и многих других пресноводных высших раков метаморфоз подавляется и все развитие зародыша протекает внутри яйце-вой оболочки. Из яйца выходит миниатюрная копия материнского организма. Развитие в этом случае становится прямым и сводится к постепенному росту организма, протекающему с помощью линек.
Линька раков, как и других членистоногих, — сложный процесс, включающий не только морфологические, но и физиологические изменения организма. Наиболее детально он изучен у высших раков. В период, предшествующий линьке, в тканях и гемолимфе линяющего животного накапливается ряд органических (липиды, белки, витамины, углеводы и т. д.) и минеральных соединений. У речного рака, например, в гиподерме откладывается гликоген, минеральные соли — в клетках печени и в особых образованиях, называемых «жерновками». Последние имеют вид округлых твердых бляшек и локализуются в стенках желудка. Частично все эти вещества поступают из старой кутикулы, которая постепенно истончается за счет растворения внутренних слоев. Возрастает интенсивность обменных процессов в организме, о чем свидетельствует усиленное потребление кислорода.
Одновременно клетками гиподермы начинает выделяться вещество новой кутикулы. Последняя постепенно утолщается, но сохраняет гибкость и эластичность. Необходимые для построения кутикулы материалы: некоторые углеводы, липиды и другие вещества — черпаются из ге-молимфы и тканей, где их содержание непосредственно перед линькой заметно возрастает, а затем постепенно снижается. Момент линьки заключается в том, что старый кутикуляриый покров лопается и через образовавшуюся щель животное выбирается наружу, оставляя пустой кутикулярный чехол — экзувий. Быстрое увеличение размеров тела у перелинявшего рака обусловлено изменениями в водном обмене, которые приводят к накоплению воды в тканях и набуханию последних. Увеличение объема тканей и органов за счет деления клеток имеет место только между линьками. Через некоторое время после сбрасывания экзувия в новых покровах откладываются минеральные соли, и кутикула быстро твердеет.
310
Весь процесс линьки находится под строгим контролем гормональной системы (см. рис. 263). Важную роль играют нейросекреторные клетки, связанные с синусовой железой (с. 303), и небольшая эндокринная железа, расположенная в головном отделе. Последняя продуцирует гормоны, запускающие и ускоряющие процесс линьки. В нейросекреторных клетках глазных стебельков образуются гормоны, подавляющие деятельность этой железы и тем самым препятствующие наступлению линьки. Особенно много этих «тормозных» гормонов вырабатывается в постлиночный и межлиночный периоды. В последующем, однако, количество их уменьшается, деятельность головной эндокринной железы активизируется и начинается подготовка к очередной линьке.
В регуляции и координации всех процессов, связанных с линькой, по-видимому, принимают участие и другие гормоны.
|
|
|
 Скачать 15 Mb.
Скачать 15 Mb.