В.А. Догель - Зоология беспозвоночных. Тип губки (spongia, или porifera)
 Скачать 15 Mb. Скачать 15 Mb.
|

|
I — мембранная трубка, или лигамент, 2 — хордотональные сенсиллы — сколпофоры. 3 — нерв, 4 — кутикула, 5 — гиподерма Типичным строением обладают тимпанальные органы кузнечиков (рис. 333, А, Б; 334), расположенные на голенях передних ног. В верхней части голени имеются по две узкие продольные щели, ведущие в два барабанных кармана. Внутренние стенки кармашков (рис. 333,£), обращенные друг к другу, тонки и представляют собой барабанные перепонки, наружные же утолщены и называются барабанными крышечками. Между обеими барабанными перепонками, вплотную примыкая к ним, проходят два трахейных ствола, которые, быть может, служат в качестве резонаторов. Наконец, главную часть тимпанального органа составляют три группы сколпофоров (рис. 334). Сколпофоры примыкают частью к барабанной перепонке, частью к резонирующей трахее. Центральные отростки чувстви-Рис 332 Внутренний тельных клеток образуют тимпанальный нерв. Точно по такому же принципу — сочетание сколпофоров и тимпанальных перепонок — устроены тимпанальные органы и других насекомых — саранчовых, сверчков, бабочек и др. Правда, располагаться они могут в разных местах тела — на передних сегментах брюшка, у основания крыльев и т. п. Хордотональные сенсиллы тимпанальных органов служат для восприятия колебаний различной частоты — имеются «высокочастотные» и «низкочастотные» сенсиллы. Как правило, одна из таких групп настроена на частоты, максимально представленные в звуках, издаваемых особями того же вида. В целом насекомые воспринимают звуки в очень широком диапазоне: от инфразвука (8—10 Гц) до ультразвука (45000 Гц). Насекомые способны не только воспринимать, но и издавать звуки. Эта особенность характерна для представителей многих групп: прямокрылых, жуков, перепончатокрылых, бабочек и др. Звуковые органы насекомых очень разнообразны. Стрекотание прямокрылых, например, вызывается развитием известных стрекочущих приспособлений, которые чаще всего связаны с крыльями. Так, у кузнечиковых эти органы находятся на передних крыльях. Некоторые жилки левого крыла становятся зазубренными и превращаются в так называемый смычок, которым животное водит по правому кры- 358 лу, где в соответствующем месте находится резонатор. Последний состо ит из ограниченной высокой жилкой площадки на крыле — зеркальца. Движение зазубренного смычка по краю зеркальца приводит к вибрации растянутой на нем части поверхности крыла. У саранчовых смычок образован рядом мельчайших зубчиков на бедрах задних ног. При трении бедер о верхние крылья зубчики задевают за сильно выдающуюся у самца радиальную жилку крыла. У самцов цикад есть своеобразный «голосовой аппарат» на нижней стороне заднегруди: действие его основано на чрезвычайно быстром колебании хитиновой перепонки, приводимой в движение сокращением мышц. Значение способности издавать звуки заключается, по-видимому, в привлечении стрекочущими самцами самок. 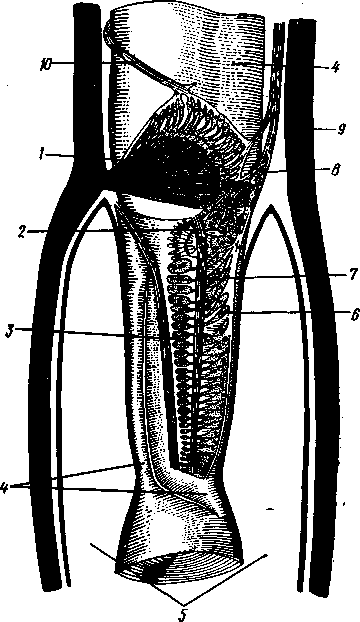 Хеморецепторы насекомых представлены обонятельными и вкусовыми сенсиллами. Кутикулярные образования обонятельных сенсилл (рис. 335, А, Б) очень разнообразны по форме: щетинки, конусовидные придатки, пластинки и т. п. Общая черта — наличие тонких пор, пронизывающих кутикулу. Через эти поры открыт доступ к чувствительным элементам сенсиллы для молекул пахучих веществ. Обонятельные сенсиллы располагаются главным образом на сяжках и челюстных щупиках. Обоняние служит насекомым как для отыскания пищи, так и при спаривании: самцы часто находят самок по запаху. Последние выделяют особые пахучие вещества — половые аттрактанты. Достаточно ничтожного количества (100 моле- 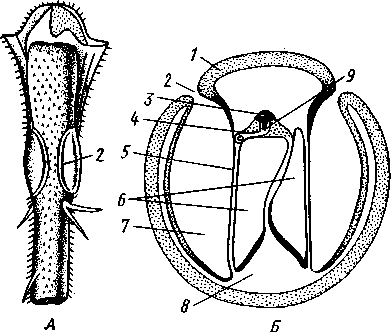 Рис. 333. Тимпанальный орган кузнечика. А — передняя голень спереди, видны два отверстия тимпанального органа (2); Б — поперечный разрез через голень в области тимпанального органа (по Швабе): / — кутикула голени, 2 — слуховая щель, 3 — шапочковая клетка, 4 — сколпофор, 5—барабанная перепонка, в — трахеи, 7— барабанная полость, 8 — полость ноги, 9 — сколопоид-ное тельце Рис. 334. Тимпанальный орган кузнечика, вскрытый с передней стороны (по Швабе): 1 — первая группа сколпофоров (подколенный орган), 2 — вторая группа сколпофоров (промежуточный орган). 3 — третья группа сколпофоров (слуховой гребень), 4 — трахейные стволы, 5 — барабанные полости, 5 — чувствительные клетки сколпофоров, 7 — сколопоид-ное тельце, * — тимпанальный нерв, 9 — кутикула, 10 — подколенный нерв 359 кул в 1 см3 воздуха) такого вещества, чтобы вызвать возбуждение у самцов шелкопряда. Вкусовые сенсиллы располагаются у насекомых на ротовых конечностях и дистальных члениках лапок. Их кутикулярные элементы представлены волосками или конусовидными придатками и также пронизаны порами. В состав каждой сенсиллы (рис. 336) входит несколько рецеп- 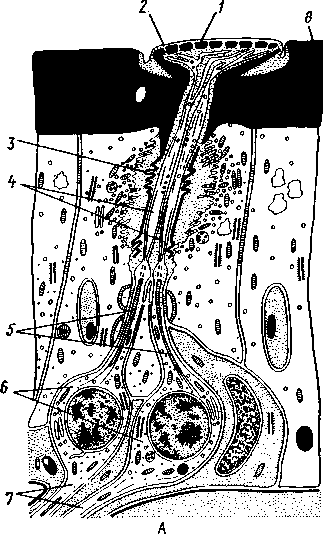 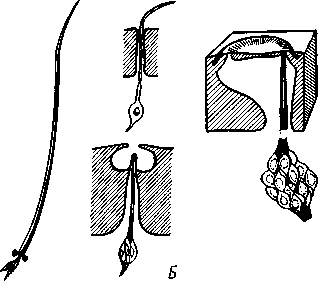 Рис. 335. Схема строения обонятельной сенсиллы насекомых. А — строение обонятельной сенсиллы (по В. Иванову); Б — различные типы обонятельных сенсилл (по Эрнсту): ; — кутикулярная дисковидная шапочка-пластинка, 2 — поры. 3 — кутикулярная трубочка, фиксирующая жгутик, 4 — видоизмененный жгутик, 5 — периферический отросток чувствительной клетки, 6 — чувствительная клетка, 7 — центральный отросток чувствительной клетки, 8—кутикула торных клеток, каждая из которых реагирует на определенный вкусовой раздражитель: одна клетка реагирует на соли, другая на сахаристые вещества, третья на чистую воду. Одна из чувствительных клеток вкусовой сенсиллы является механорецепторной. Таким образом, у насекомых, так же как и у позвоночных, вкусовое ощущение сопровождается осязательным. Наиболее сложными из органов чувств у насекомых являются органы зрения. Последние представлены образованиями нескольких типов, из которых важнейшие — сложные фасетированные глаза примерно такого же строения, как и сложные глаза ракообразных. Глаза состоят из отдельных омматидиев (рис. 337), количество которых определяется главным образом биологическими особенностями на- \ секомых. Активные хищники и хорошие летуны, стрекозы обладают! глазами, насчитывающими до 28 000 фасеток в каждом. В то же время \ муравьи (отр. Перепончатокрылые), особенно рабочие особи видов, обитающих под землей, имеют глаза, состоящие из 8—9 омматидиев. 360 Каждый омматидий представляет совершенную фотооптическую сенсиллу (рис. 338, А, Б).В его состав входят оптический аппарат, включающий роговицу, — прозрачный участок кутикулы над омматидием и так называемый хрустальный конус. В совокупности они выполняют роль линзы. Воспринимающий аппарат омматидия представлен несколь- 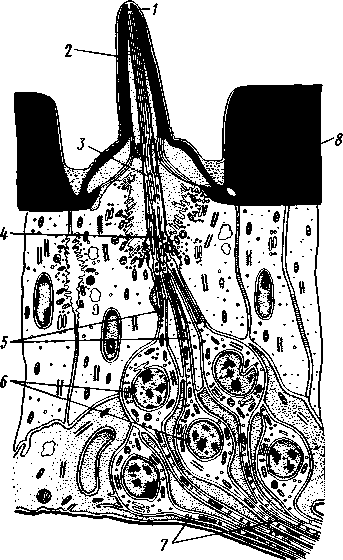 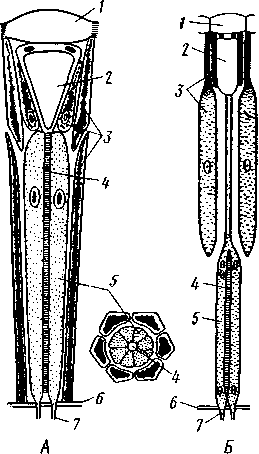 Рис. 336. Схема строения вкусовой сенсиллы насекомых (по В. Иванову): 1 — пора, 2 — кутикулярный конус, 3 — кутику-лярная трубочка, фиксирующая жгутик, 4 — видоизмененный жгутик, 5—периферический отросток чувствительной клетки, 6 — чувствительная клетка, 7 — центральный отросток чувствительной клетки, 8 — кутикула Рис. 338. Схема строения омма-тидиев дневных насекомых (А) и ночных и сумеречных насекомых (Б) (из Мазохина-Порш-някова): / — прозрачный участок роговицы — хрусталик, 2 — хрустальный конус, 3 — пигментные клетки, 4 — рабдом. 5 — чувствительные клетки, 6 — ба-зальная мембрана, 7 — центральные отростки чувствительных клеток 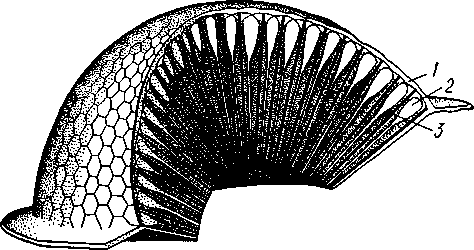 Рис. 337. Схема строения сложного глаза насекомого. Участок глаза вырезан (по Гессе): / — роговица — прозрачная кутикула. ? — xpvc-тяльный конус. 3 — пигмент между омматиди-ями 361 кими (4—12) рецепторными клетками; специализация их зашла очень далеко, о чем говорит полная утрата ими жгутиковых структур. Собственно чувствительные части клеток — рабдомеры — представляют скопления плотно упакованных микроворсинок, располагаются в центре омматидия и тесно прилегают друг к другу. В совокупности они образуют светочувствительный элемент глаза — рабдом. По краям омматидия залегают экранирующие пигментные клетки; последние довольно существенно отличаются у дневных и ночных насекомых. В первом случае пигмент в клетке неподвижен и постоянно разделяет соседние омматидии, не пропуская световые лучи из одного глазка в другой. Во втором случае пигмент способен перемещаться в клетках и скапливаться только в их верхней части. При этом лучи света попадают на чувствительные клетки не одного, а нескольких соседних омматидиев, что заметно (почти на два порядка) повышает общую чувствительность глаза. Естественно, что подобного рода адаптация возникла у сумеречных и ночных насекомых. От чувствительных клеток омматидия отходят нервные окончания, образующие зрительный нерв. Кроме сложных глаз многие насекомые имеют еще и простые глазки (рис. 339), строение которых не соответствует строению одного омматидия. Светопреломляющий аппарат линзообразной формы, сразу же под ним расположен слой чувствительных клеток. Весь глазок одет чехлом из пигментных клеток. Оптические свойства простых глазков таковы, что воспринимать изображения предметов они не могут. Личинки насекомых в большинстве случаев обладают только простыми глазками, отличающимися, однако, по строению от простых глазков взрослых стадий. Никакой преемственности между глазками взрослых особей и личинок не существует. Во время метаморфоза (с. 373) глаза личинок полностью резорбируются. Зрительные способности насекомых совершенны. Однако структурные особенности сложного глаза предопределяют особый физиологический механизм зрения. Животные, имеющие сложные глаза, обладают «мозаичным» зрением. Малые размеры омматидиев и их обособленность друг от друга приводят к тому, что каждая группа чувствительных клеток воспринимает лишь небольшой и сравнительно узкий пучок лучей. Лучи, падающие под значительным углом, поглощаются экранирующими пигментными клетками и не достигают светочувствительных элементов омматидиев. Таким образом, схематично каждый омматидии получает изображение только одной небольшой точки объекта, находящегося в поле зрения всего глаза. Вследствие этого изображение складывается из стольких световых точек, отвечающих различным частям объекта, на сколько фасеток падают перпендикулярно лучи от объекта. Общая картина комбинируется как бы из множества мелких частичных изображений путем приложения их одного к другому. Восприятие.цвета насекомыми также отличается известным своеобразием. Представители высших групп Insecta имеют цветовое зрение, основанное на восприятии трех основных цветов, смешение которых и дает все красочное многообразие окружающего нас мира. Однако у насекомых по сравнению с человеком наблюдается сильный сдвиг в коротковолновую часть спектра: они воспринимают зелено-желтые, синие и ультрафиолетовые лучи. Последние для нас невидимы. Следовательно, цветовое восприятие мира насекомыми резко отличается от нашего. Функции простых глазков взрослых насекомых требуют еще серьезного изучения. По-видимому, они в какой-то мере «дополняют» сложные глаза, влияя на активность поведения насекомых в разных условиях ос- 362 вещенности. Кроме того, было показано, что простые глазки наряду со сложными глазами способны воспринимать поляризованный свет. Помимо перечисленных органов чувств насекомые обладают еще рядом рецепторных аппаратов. Таковы сенсиллы, воспринимающие температуру окружающей среды, ее влажность. Водные насекомые способны регистрировать изменения давления и т. п. Органы дыхания. Для дыхания служит сложно развитая система трахей (рис. 340). По бокам тела находится до 10 пар, иногда меньше, дыхалец, или стигм: они лежат на средне- и заднегруди и на 8 члениках брюшка. 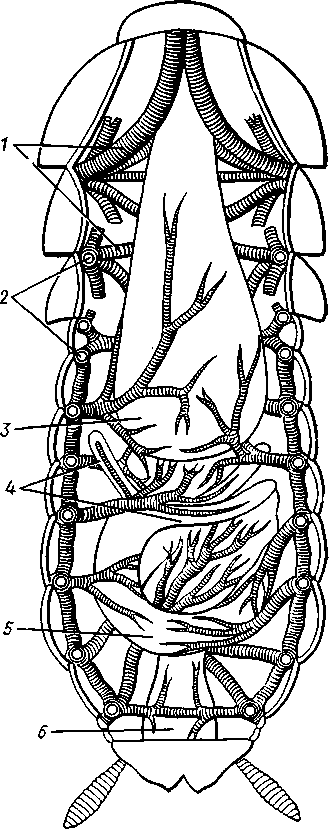 Стигмы часто снабжены особыми замыкательными аппаратами и ведут каждая в короткий поперечный канал, а все поперечные каналы соединены между собой парой (или больше) главных продольных трахейных стволов. От стволов берут начало более тонкие трахеи, ветвящиеся многократно и опутывающие своими разветвлениями все органы. Заканчивается каждая трахея концевой клеткой с радиально расходящимися отростками, пронизанными конечными канальцами трахеи (рис. 341). Концевые веточки этой клетки (трахеолы) проникают даже внутрь отдельных клеток тела. Иногда трахеи образуют местные расширения, или воздушные мешки, которые служат у наземных насекомых для улучшения вентиляции воздуха в трахейной системе, а у вод- 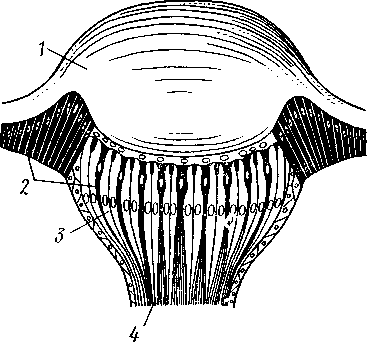 Рис. 339. Схема строения простого глазкя (по Веберу): / — прозрачный участок роговицы — хрусталик, 2 — пигментные клетки, 3 — чувствительные клетки, 4 — центральные отростки чувствительных клеток Рис. 340. Трахейная система черного таракана со спинной стороны (по Майаль и Денни): 1 — трахейные стволы. 2 — стигмы. 3 — зоб, 4 — пилорические придатки. 5 — средняя кишка. 6 — задняя кишка 363 o> 4» 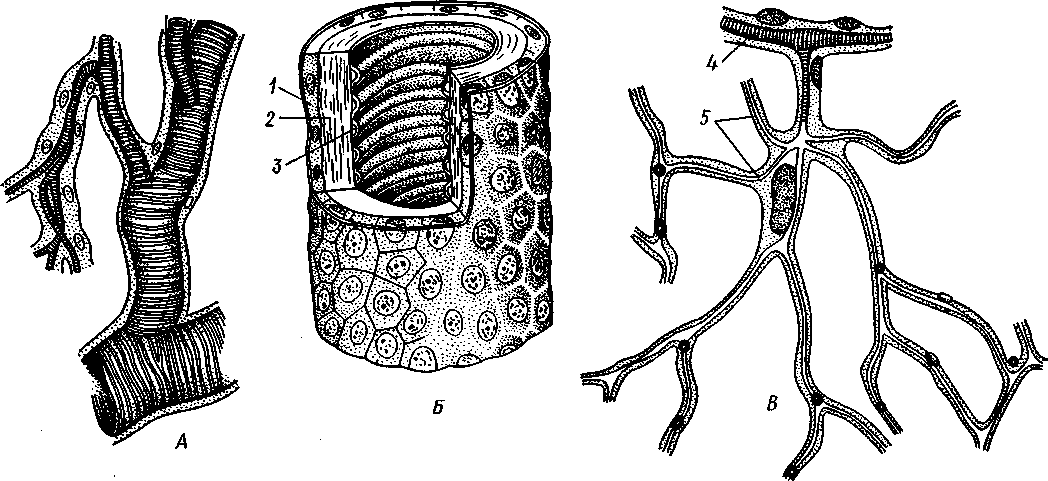 Рис. 341. Строение трахей насекомых. А — участок трахеи с разветвлениями; Б — стенка трахеи при большом увеличении; В — концевая клетка трахеи (по Веберу и Гольмгрену): / — эпителий трахеи, 2 — хитиновая выстилка трахеи, 3 —спиральная нить — утолщение хитиновой выстилки трахеи, 4 — трахея, 5 — тра- хеолы (тончайшие ответвления без хитиновой выстилки) ных, вероятно, в качестве резервуаров, увеличивающих запас воздуха в теле животного. Трахеи возникают у зародыша насекомых в виде глубоких впячиваний эктодермы; как и остальные эктодермальные образования, они выстланы кутикулой (рис. 341,5). В поверхностном слое последней образуется спиральное утолщение, придающее трахее эластичность и препятствующее спадению стенок. В простейших случаях поступление кислорода в трахейную систему и удаление из нее углекислого газа происходит путем диффузии через постоянно открытые стигмы. Это наблюдается, однако, только у малоактивных насекомых, обитающих в условиях повышенной влажности.  Активизация поведения и переход к обитанию в засушливых биотопах значительно усложняют механизм дыхания. Возрастающая потребность организма в кислороде обеспечивается появлением специальных дыхательных движений, состоящих из расслабления и сжимания брюшка. При этом происходит вентилирование трахейных мешков и основных трахейных стволов. Образование замыкательных аппаратов на стигмах снижает потери воды в процессе дыхания. Так как скорость диффузии паров воды ниже, чем кислорода, то при кратковременном открывании стигм кислород успевает проникнуть в трахейную систему, а потери воды оказываются минимальными. У многих живущих в воде личинок насекомых (например, стрекоз, поденок и др.) трахейная система является замкнутой, т. е. стигмы отсутствуют, тогда как сама трахейная сеть налицо. У таких форм кислород диффундирует из воды через трахейные жабры, пластинчатые или кустистые, тонкостенные выросты тела, пронизанные богатой сетью трахей (рис. 342). Чаще всего трахейные жабры сидят по бокам части члеников брюшка (личинки поденок). Кислород поступает через тонкие покровы жабр, попадает в трахеи и затем разносится по телу. Во время превращения жабродышащих личинок во взрослое насекомое, живущее на суше, жабры исчезают, а стигмы открываются и трахейная система из замкнутой переходит в открытую. 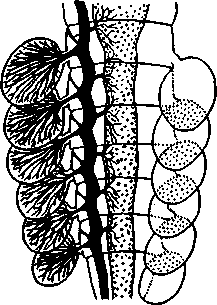 Рис. 342. Листо видные трахейные жабры личинки по денки (трахеи за чернены) (из Шванвича) Рис. 343. Строение сердца жука-плавунца Dyti-scus marginalis (из Кузнецова) : / — аорта, 2 — сердце. 3 — остии, 4 — крыловидные мышцы 365 Важная физиологическая особенность дыхательной системы насекомых состоит в следующем. Обыкновенно кислород воспринимается животным в определенных участках его тела и оттуда разносится кровью по всему организму. У насекомых же воздухоносные трубочки пронизывают все тело и доставляют кислород прямо к местам его потребления, т. е. к тканям и клеткам, как бы заменяя собой кровеносные сосуды. Кровеносная система в связи с отмеченной особенностью дыхательной системы развита у насекомых сравнительно слабо. В брюшке над кишечником залегает длинное трубковидное сердце (рис. 343). Задний конец его слепо замкнут, а полость поделена перегородками, несущими клапанные отверстия, на несколько камер (чаще 8). В стенках сердца залегают мышечные волокна, обеспечивающие его сокращение. Каждая камера снабжена парой боковых остий. На переднем конце сердце продолжается в мускулистую головную аорту, которая, достигнув мозга, 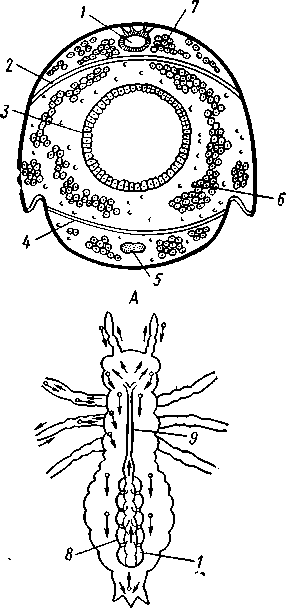 заканчивается отверстием, так что гемо-лимфа из нее поступает прямо в полость тела. |