В.А. Догель - Зоология беспозвоночных. Тип губки (spongia, или porifera)
 Скачать 15 Mb. Скачать 15 Mb.
|

|
I — верхняя губа, 2—верхняя челюсть—мандибула, .3—нижнечелюстной щупик, 4 — наружная лопасть нижней челюсти, 5 — внутренняя лопасть нижней челюсти, 6 — нижняя челюсть, состоящая из стволика и основного членика, 7 — нижняя губа, состоящая из подбородка и подподбородка, 8 — внутренняя лопасть нижней губы, 9 —• В состав ротового аппарата грызущего типа (рис. 310) входят следующие образования. Спереди рот прикрыт «верхней губой», представляющей складку покровов головы и образующей переднюю стенку ротовой полости. Верхняя губа закладывается независимо от конечностей и не гомологична им. За ней следует пара верхних челюстей— жвал, или мандибул. Это две толстые нерасчлененные пластинки, зазубренные по внутреннему краю и играющие наиболее важную роль в размельчении пищи. Мандибулы являются конечностями II сегмента головы. Конечности III и IV сегментов — максиллы, или нижние челюсти. Они члени-сты н этим напоминают О происхождении ОТ ходных ног. Первая пара нижних челюстей г г раСПОЛОЖеНЗ ПО бОКЗМ рТЗ, ПрИЧбМ КЭЖДЗЯ челюсть СОСТОИТ ИЗ ДВуЧЛбНИКОВОГО ОСНОВа- ния и сидящих на его вершине трех придатков: двух лопастей, внутренней и наруж- НОЙ- И ЧЛСНИСТОГО Ч6ЛЮСТНОГО ЩупИКЗ. ЭтО НЗИбоЛСС богЗТО раСЧЛеНбННЫе КОНСЧНОСТИ насекомых, вследствие чего как раз на строении первой пары максилл базируются попытки провести гомологию между конечностями Insecta и основным двуветвистым типом конечностей другой ветви членистоногих, а именно ракообразных. Вторая пара нижних челюстей насекомых сливается (как у многоножек Chilopoda, см. рис. 302) и образует непарную пластинку — нижнюю губу. Соответственно происхождению нижняя губа состоит из основной пластинки и сидящих на ней трех пар придатков. Последние представляют собой две пары нерасчлененных лопастей и пару нижнегубных щупиков. К ротовому аппарату относится также хитиновое выпячивание дна ротовой полости — гипофаринкс, или язык. Наименьшее сравнительно с описанным основным типом изменение обнаруживают лакающие ротовые органы (рис. 311) многих перепончатокрылых (пчелы, шмели). Верхняя губа и жвалы приблизительно такого же строения, как и в грызущем ротовом аппарате. Напротив, обе пары нижних челюстей заметно изменяются по сравнению с исходным типом: сохраняя полный набор частей, входящих в их состав, они сильно вытягиваются в длину и в сложенном «рабочем» состоянии образуют довольно широкий хоботок. Подобная двойственность в строении ротового аппарата пчел и шмелей объясняется особенностями их питания. Мандибулы служат для сбора и размалывания твердой цветочной пыльцы, а хоботок, образуемый максиллами, — для всасывания нектара. Переход многих насекомых к питанию только жидкой пищей привел к возникновению более специализированных ротовых аппаратов, 342 приспособленных для сосания, основу которых составляет хорошо герметизированная трубка. Последняя в разных отрядах насекомых имеет различное происхождение и строение в зависимости от особенностей источника и характера пищи. У комаров (отр. Diptera), например, совокупность всех ротовых частей образует колющий хоботок (рис. 312), состоящий из желобовидного футляра, в котором заложены колющие щетинки. Желоб образован очень сильно вытянутой нижней губой, щупики которой почти полностью атрофируются. Сверху желоб прикрыт тоже вытянутой верхней губой, края которой смыкаются, образуя узкую трубку, служащую для всасывания крови. Жвалы, две пары нижних челюстей и гипофаринкс преобразованы в тонкие колющие стилеты, легко проникающие через покровы позвоночных животных. 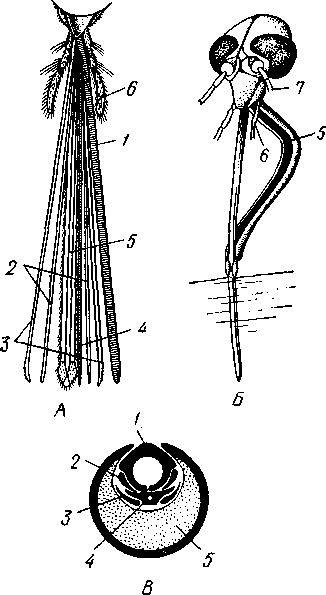 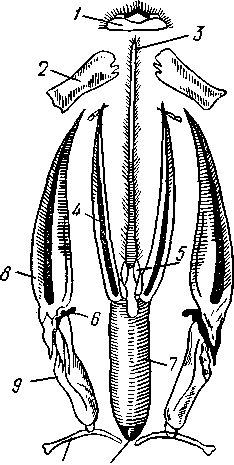 Ю 11 Сосущие ротовые органы (рис. 313) встречаются у чешуекрылых и устроены в виде сосательного хоботка. Ротовой аппарат характеризуется необычайным развитием первой пары нижних челюстей и редукцией всех остальных частей. Верхняя губа едва различима, жвал нет. Нижняя губа образует небольшую нерасчлененную треугольную пластинку, снабженную трехчлениковыми щупиками. Нижняя челюсть каждой Рис. 311. Лакающие ротовые органы шмеля Bombus (по Богданову-Катькову): / — верхняя губа, 2 — верхняя челюсть, 3 — язычок — сросшиеся внутренние лопасти нижней губы, 4 — нижнегубной щупик, 5 — наружная лопасть нижней губы, 6 — нижнечелюстной щупик, 7 — подбородок, 5 — нижняя челюсть. 9 — стволик, 1П — основной членик, // — подпод-бородок Рис. 312. Колющие ротовые органы комара. А — хоботок комара в расправленном виде (по Муру); Б — положение ротовых частей во время акта сосания крови (по Веберу); В — поперечный разрез через хоботок комара (из Зеликмана): / — верхняя губа, 2 — мандибула. 3 — нижняя челюсть, 4 — гипофаринкс, 5—нижняя губа, 6 — «ижне-челюстной щупик, 7 — сяжки 343 стороны вытянута в очень длинный желобок, вогнутость которого обращена к медианной линии тела. Желобки обеих сторон плотно прикладываются друг к другу краями, образуя трубку. В покоящемся состоянии хоботок свернут на брюшной стороне в крутую спираль и спрятан под 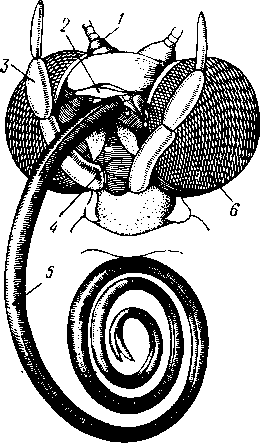 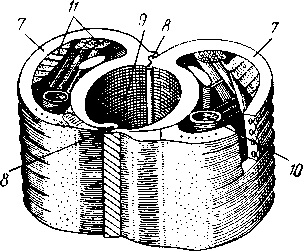 Рис. 313. Сосущий ротовой аппарат бабочки. /1—голова бабочки с расправленным хоботком; Б — участок хоботка при большом увеличении (по Веберу): / — основание усиков, 2 — верхняя губа, 3 — нижнегубной щупик, 4 — нижняя губа, 5 — хоботок — нижние челюсти, 6 — фасетированный глаз, 7 — правая и левая нижние челюсти, S — места соединения челюстей, 9 — полость хоботка, 10 — трахеи, 11 — мускулатура хоботка головой. В расправленном виде хоботок засовывается внутрь цветков при высасывании нектара. Лижущим ротовым аппаратом (рис. 314) обладают многие мухи из двукрылых. Главная часть аппарата — мясистая нижняя губа (хоботок), заканчивающаяся двумя большими пластинчатыми выростами, последние снабжены сложно устроенным фильтрующим аппаратом. Мандибулы и первая пара нижних челюстей атрофированы, хотя челюстные щупики сохраняются. Верхняя губа и гипофаринкс, расположенные в желобооб-разном углублении на передней стенке нижней губы, в совокупности с ней образуют трубочку, в которую и поступает жидкая пища, слизанная и профильтрованная пластинчатыми выростами нижней губы. В некоторых случаях может происходить вторичное упрощение или даже почти полная редукция ротового аппарата. Это главным образом характерно для форм, не питающихся на взрослой стадии (поденки, оводы). Описанные крайние ступени развития ротового аппарата связаны с основным грызущим типом ротовых конечностей рядом переходов. Положение ротовых частей на головной капсуле имеет важное систематическое значение. У подавляющего большинства форм они расположены снаружи, на поверхности головы. Эти насекомые объединяются в подкласс Открыточелюстных (Ectognatha). У небольшого числа самых примитивных форм ротовые конечности погружены внутрь особой рото- 344 вой капсулы, так что наружу в лучшем случае торчат только их кончики. Подобные насекомые составляют второй подкласс — Скрыточелюстные (Entognatha). Грудь насекомых (см. рис. 307) состоит из трех сегментов, называемых передне-, средне- и заднегрудыо. Сегменты груди несут три пары 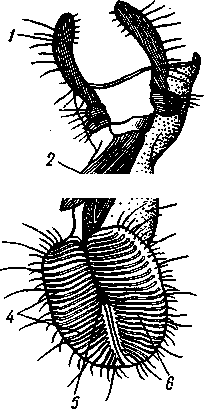 двигательных конечностей, прикрепляющихся между стернитом и боковой пластинкой каждой стороны (рис. 315). Конечности всегда состоят из одного ряда члеников, в которых мы отличаем от основания к концу ноги: 1) тазик, или ляжку, широкий основной членик; 2) вертлуг; 3) бедро, самый толстый членик ноги; 4) голень, обычно самый длинный из члеников; з^шгт _ 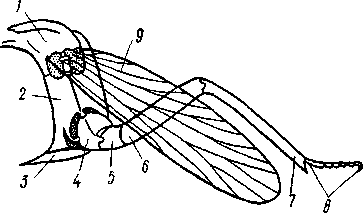 7 Рис. 314. Лижущий ротовой аппарат мухи Musca domestica. Вид спереди и снизу (из Шван-вича); 1 — нижнечелюстной щупик, 2 — верхняя губа, 3 — гипофаринкс, А — каналы фильтрующего аппарата. 5 — ротовое отверстие, 6 — лопасти нижней губы, 7 — нижняя губа Рис. 315. Схема строения грудного сегмента (по Шванвичу): / — тсргит, 2 — боковая пластинка, 3 — стернит. 4 —тазик, 5 — вертлуг. f — бедро, 7 — голень, 8 — лапка, 9 — крыло 5) лапку, состоящую из разного числа (максимально до 5) очень маленьких члеников. Последний из них несет на конце два (реже один) коготка. В связи с разными способами движения ноги испытывают модификации,иногда очень сильные (рис. 316). Ближе всего подходят к 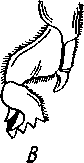 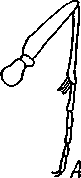 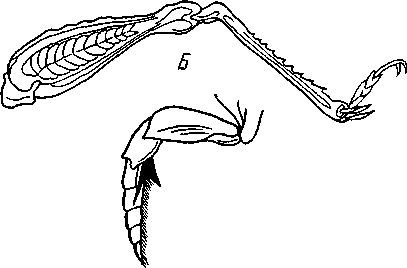 Г 345 Рис. 316. Различные типы конечностей насекомых. А—бегательная; Б — пры-гательная; В — копательная; Г — плавательная (из Бей-Биенко) описанному типу бегательные ноги, которые и наиболее распространены у Insecta. У насекомых с прыгательными ногами, например у кузнечиков, бедро и голень задней пары ног сильно вытягиваются. У роющих насекомых все ноги, а в особенности передние, играющие главную роль при копании, укорачиваются, становятся массивными и приобретают мощное вооружение из хитиновых зубцов. Плавательные конечности сплющены в виде весла и снабжены густым рядом упругих гребных волосков (жуки-плавунцы). Конечности насекомых, представляющие систему подвижно соединенных друг с другом рычагов с большим числом степеней свободы, способны к разнообразным и совершенным движениям. Характернейшей особенностью насекомых как большой систематической группы является их способность к полету. Полет осуществляется 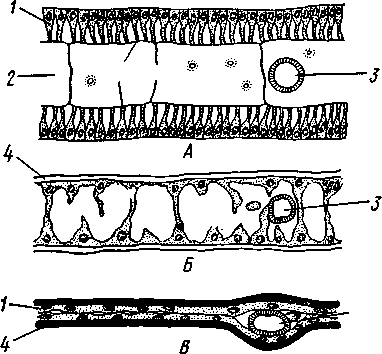 7 2 J 4 ff 1 -3 Рис. 317. Последовательные стадии формирования крыла бабочки. А — зачаток крыла куколки; Б — крыло только что вышедшей из куколки бабочки; В — затвердевшее крыло (из Шванвича): / — гиподерма, 2 — полость зачатка крыла, 3 — трахея, 4—кутикула Рис. 318. Схема жилкования крыла насеко- ] мого (из Кенигсмена): / — костальная жилка, 2 — субкостальная жилка, 3— радиальная жилка, 4 — медиальная жилка, I 5 — кубитальная жилка, 6 — анальные жилки, 7 — югальные жилки при помощи крыльев; в большинстве случаев их две пары и расположены они на II (среднегрудь) и III (заднегрудь) грудных сегментах. Крылья представляют, по существу, мощные складки стенки тела. Хотя полностью сформированное крыло имеет вид тонкой цельной пластинки, оно тем не менее двухслойно; верхний и нижний слои разделены тончайшей щелью, являющейся продолжением полости тела (рис. 317,В). В крыло, как и во все участки тела, заходят трахейные стволы и нервы. В местах их залегания на крыльях образуются характерные трубчатые утолщения — жилки, располагающиеся строго определенным для каждого вида образом, в результате чего возникает определенный рисунок— жилкование. В настоящее время выработана единая система названий жилок, а исходный тип жилкования представляется следующим образом (рис. 318). Вдоль крыла проходят так называемые продольные жилки: костальная, субкостальная, радиальная, медиальная, кубитальная, анальные и югальные. Все они, кроме первой, могут образовывать дополнительные ветви. Наряду с продольными имеются и поперечные жилки. 346 Особенности строения крыльев становятся хорошо понятными при знакомстве с процессом их развития в онтогенезе насекомых (см. рис. 317). Крылья закладываются в виде мешкообразных выпячиваний кожи, в которые продолжаются полость тела и трахеи. Выпячивания сплющиваются дорзовентрально; гемолимфа из них оттекает внутрь тела, верх- 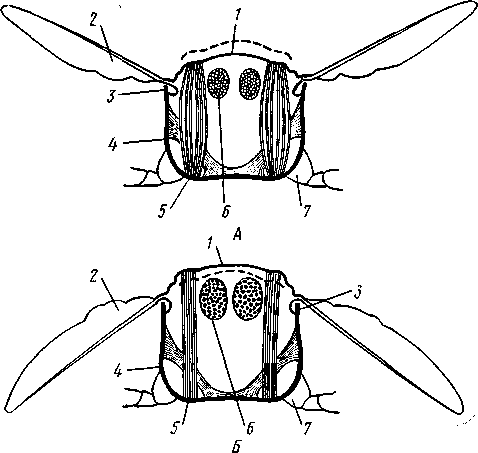 Рис. 319. Схема работы крыльев насекомого. А —стадия поднятого крыла; Б — стадия опущенного крыла (по Шванвичу): I — тергит, 2 — крыло, 3 — столбик, 4 — боковая пластинка, 5 — дорзо-вентральная мышца, 6 — продольная мышца, 7 — основание конечности ний и нижний листки пластинки сближаются, мягкие ткани частично дегенерируют, и крыло приобретает вид тонкой перепонки. Движение крыльев у насекомых — результат работы сложного механизма и определяется, с однойстороны, особенностью сочленения крыла с туловищем, а с другой — действием особых крыловых мышц. В общих чертах основной механизм движения крыльев представляется следующим образом (рис. 319). Само крыло — это двуплечий рычаг с неодинаковой длиной плечей. С тергитом и боковой пластинкой крыло соединяется тонкими и гибкими мембранами. Чуть отступя от места этого соединения, крыло опирается на небольшой, имеющий вид столбика вырост боковой пластинки, который и является точкой опоры крылового рычага. Расположенные в грудных сегментах мощные продольные и дорзо-вентральные мышцы могут опускать или приподнимать тергит. При опускании последний давит на короткое плечо крыла и влечет его за собой вниз. В результате длинное плечо, т. е. вся несущая плоскость крыла, движется вверх. Подъем тергита приводит к опусканию крыловой пластинки (рис. 319). Небольшие мышцы, прикрепленные непосредственно к крылу, способны поворачивать его вдоль продольной оси, при этом изменяется угол атаки. Во время полета свободный конец крыла движется по довольно сложной траектории (рис. 320). При опускании крыловая 347 пластинка расположена горизонтально и движется вниз и вперед: возникает подъемная сила, удерживающая насекомое в воздухе. При движении вверх и назад крыло располагается вертикально, что создает про-пеллирующий эффект. Количество ударов крыла в 1 с сильно варьирует у разных насекомых: от 5—10 (у крупных дневных бабочек) до 500—600 (многие комары); у 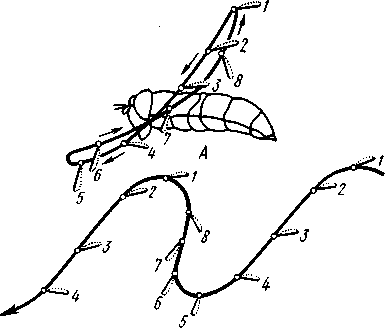 Рис. 320. Схема траектории крыла насекомого. А — при стоячем полете мухи-журчалки; Б — при полете вперед (по Залесскому): 1—8 — восемь последовательных стадий положений крыла (сплошная черта — нижняя сторона крыла, пунктир — верхняя сторона) очень мелких комаров-мокрецов эта цифра достигает 1000 колебаний в 1 с. У различных представителей насекомых передние и задние крылья могут быть развиты в разной степени. Только у более примитивных насекомых (стрекозы) обе пары крыльев развиты более или менее одинаково, хотя и разнятся по форме. У жуков (отр. Жесткокрылые — Coleo-ptera) передние крылья изменяются в толстые и твердые надкрылья — элитры, которые почти не участвуют в полете и в основном служат для защиты спинной стороны тела. Настоящими же крыльями являются лишь задние крылья, которые в покоящемся состоянии спрятаны под надкрыльями. У представителей отряда клопов твердеет лишь основная половина передней пары крыльев, вследствие чего эту группу насекомых нередко называют отрядом Полужесткокрылых. У некоторых насекомых, а именно у целого отряда двукрылых, развита лишь передняя пара крыльев, тогда как от задней остаются лишь рудименты в виде так называемых жужжалец. 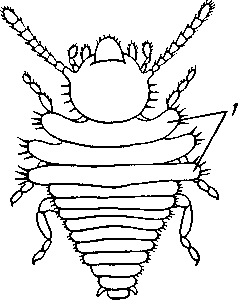 Среди насекомых имеются и бескрылые, которые делятся на две различные группы. У одних отсутствие крыльев есть черта, присущая им искони и указывающая на примитивность их организации. Это все представители подкласса Entognatha и наиболее примитивные из открыточелюст-ных — отряд Thysanura (см. рис. 366). Среди остальных Ectognatha тоже встречаются формы, лишенные крыльев, — вши, блохи, некоторые мухи и т. п. Однако бескрылость в этом случае — свойство вторичное: это формы, утратившие крылья вследствие паразитического образа жизни или каких-либо иных причин. Рис. 321. Нимфа Calotermes dilatatus с крыловидными расширениями (1)на всех сегментах груди (из Иммса) Вопрос о происхождении крыльев еще не вполне разрешен. В настоящее время одной из наиболее обоснованных представляется «паранотальная» гипотеза, согласно которой крылья возникли из простых неподвижных боковых выростов кожи — паранотумов. Такие выросты встречаются у многих члени- |