учебник. Учебник ФЛ - Виноградов, Каткова 2016. Учебник для медицинских и фармацевтических учреждений среднего профессионального образования Под
 Скачать 5.41 Mb. Скачать 5.41 Mb.
|

Средства, действующие на передачу импульсов в адренергических синапсахОсновным медиатором адренергических волокон является норадреналин. Хи- мически близкий к нему адреналин вырабатывается клетками мозгового вещества надпочечников и выделяется ими в кровь, т. е. является гормоном. Реже в окон- чаниях адренергических нервов роль медиатора выполняет дофамин. Эти три вещества близки не только по химическому строению (они являются катехола- минами), но и по фармакологическому действию. Передача импульсов в адренер- гических синапсах подчиняется рассмотренным выше закономерностям, отличия касаются в основном химической стороны процесса. Особенности адренергической передачи импульсов. Исходным продуктом для синтеза катехоламинов (КА) является аминокислота тирозин, в свою очередь тирозин может образовываться в печени из фенилаланина. До стадии дофамина синтез идет в цитозоле, затем дофамин активно поглощается везикулами (рис. 13), которые содержат необходимые ферменты. При помощи них происходит даль- нейшее превращение дофамина. В норадренергических синапсах из дофамина об- разуется норадреналин, в адренергических (нейроны некоторых областей ЦНС, мозговое вещество надпочечников) — адреналин. В дофаминергических синапсах синтез останавливается на дофамине, который и является медиатором. Депонирование КА в везикулах происходит за счет связывания со специфиче- ским белком и АТФ. Различают три формы КА в нервных окончаниях: резервный пул в везикулах (до 80 % депонированного КА), который не освобождается при поступлении нервного импульса до истощения осталь- ных пулов; мобилизационный пул II (остальные 20 %) — непосредственно выбрасыва- ется в синаптическую щель при поступлении импульса; мобилизационный пул I (до 30—40 % общего содержания катехоламинов в окончании) — медиатор, реабсорбированный из синаптической щели («отрабо- тавший»), и избыточный медиатор, который не депонирован в везикулах вслед- ствие их насыщения; КА легко вытесняются из последнего пула при введении адреномиметиков непрямого действия — эфедрина, мезокарба и др. Между тремя пулами КА существует динамическое равновесие. Освобождение КА окончаниями волокон происходит так же, как и ацетилхолина. Гормон моз- гового вещества надпочечников — адреналин — приносится в органы с кровью и поглощается клетками. Он также депонируется в нервных окончаниях и вместе с норадреналином может выделяться в синаптическую щель. Инактивация выделившихся в синаптическую щель КА идет по двум путям: внутриклеточное окислительное дезаминирование с помощью моно- аминоксидазы (МАО) митохондрий (таким способом инактивируется 10—20 % медиатора); МАО имеется также в печени и других тканях и обеспечивает инакти- вацию порций медиатора, диффундирующих из синапса и секретируемых надпо- чечниками катехоламинов; метилирование с помощью особого фермента, встроенного в постсинапти- ческие мембраны, в мембраны печеночных и других клеток,— катехол-орто-ме- тилтрансферазы (КОМТ) (по этому пути инактивации идет до 80—90 % метабо- лизирующихся КА). В целом инактивируется очень небольшая часть выделившихся в синаптиче- скую щель КА, большая часть (около 80 %) быстро подвергается обратному за- хвату («реаптейк») пресинаптическими окончаниями и повторно используется в синаптической передаче. Взаимодействие КА с рецепторами имеет ряд важных особенностей, знать ко- торые необходимо для правильного выбора и применения большой группы фар- макологических веществ. Существование трех катехоламиновых медиаторов — норадреналина, дофамина и адреналина — эволюционно обусловлено и не явля- ется случайным. Каждый из них обладает преобладающим сродством к тому или иному типу рецепторов, благодаря чему нервная система может более дифферен- цированно влиять на функции органов. 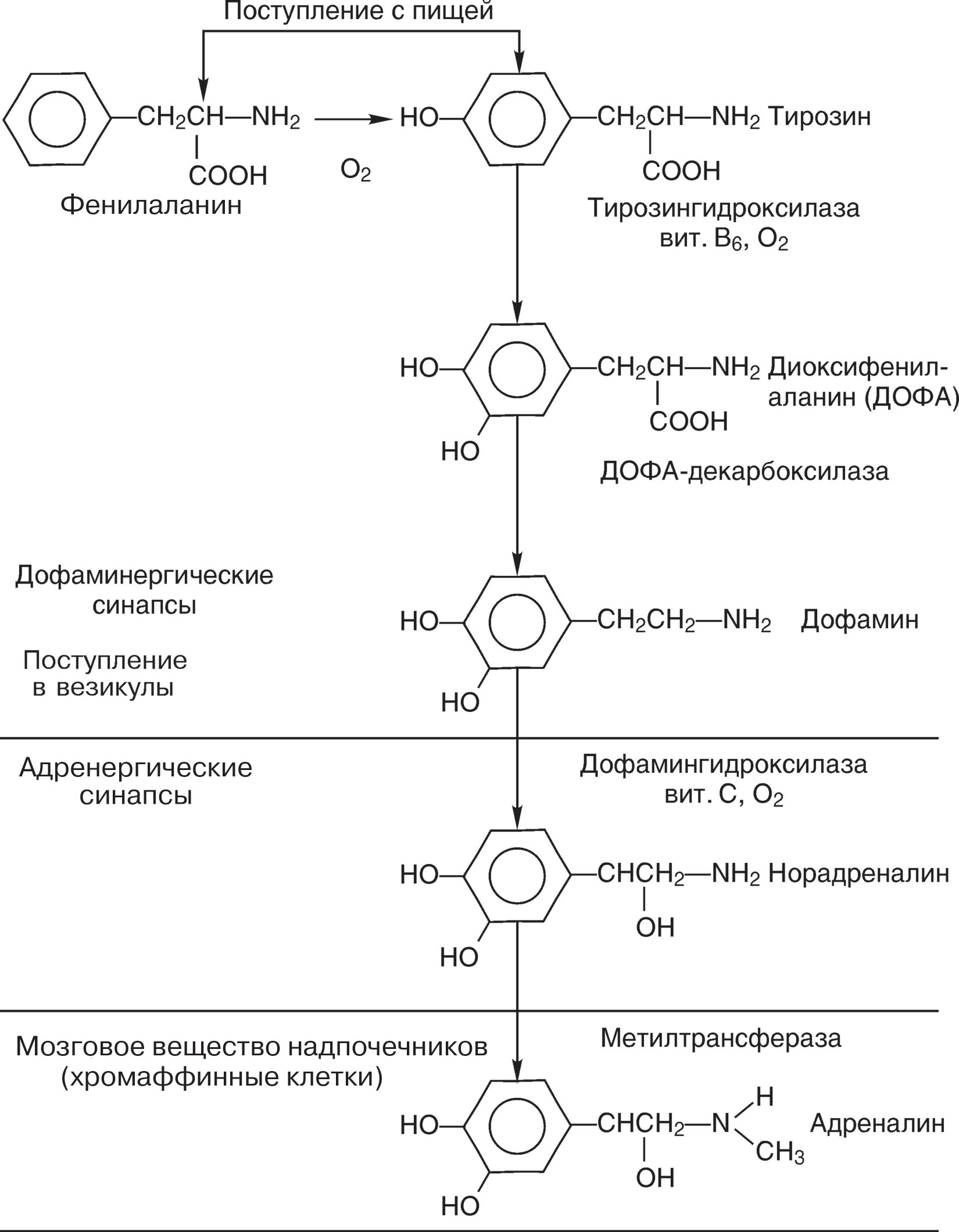 Рис.13.Стадии биосинтеза катехоламинов Работами Р. Алквиста (1948) и других фармакологов установлено, что адрено- рецепторы можно разделить на два типа (альфа и бета — - и -). В свою очередь каждый тип подразделяется на подтипы: альфа1 и альфа2; бета1 и бета2 (в жиро- вых клетках выделяют бета3-адренорецепторы). Эти рецепторы различаются по преобладающей локализации в организме, имеют различную плотность в тканях, различное функциональное значение. Дофаминовые рецепторы также подраз- деляются на несколько подтипов с разной локализацией и функцией. В синапсах адренорецепторы и дофаминовые рецепторы находятся как на постсинаптической мембране, так и на пресинаптической. Адренорецепторы могут располагаться и вне синапсов (внесинаптические или «свободные» адренорецепторы) (табл. 31). Таблица31 Локализация в организме и функциональное значение адренорецепторов
Окончаниетабл.31
* Адренорецепторы жировых клеток относят к подтипу бета3, через них активируется липолиз. Примечание. В таблицу включены наиболее важные по своей функциональной значимости адре- норецепторы. Сопряжение альфа- и бета-адренорецепторов с функциями клеток осущест- вляется посредством активации или ингибирования тех же внутриклеточных ме- ханизмов, которые были описаны в главе, посвященной холинорецепторам. Аль- фа1-адренорецепторы (RIII на рис. 11), активируя (через Gq-белки) фосфолипазу С, стимулируют образование ИТФ и ДАГ, способствуют выходу кальция из внутри- клеточных депо и активации соответствующих протеинкиназ, которые в числе прочих эффектов вызывают сокращение гладких мышц. Альфа2-адренорецепторы ингибируют аденилатциклазу (через Gi-белок) и уменьшают образование цАМФ в клетках (RII на рис. 11), при этом снижается активность цАМФ-зависимых про- теинкиназ; некоторые эффекты (торможение выброса медиаторов из пресинапти- ческих окончаний) связаны также с усилением выхода калия, гиперполяризацией мембраны и затруднением активации кальциевых каналов. Механизмы сопряжения бета-адренорецепторов с функциями клеток изучены лучше. Бета1- и бета2-адренорецепторы через Gs-белок активируют аденилатци- клазу и интенсифицируют образование цАМФ в клетках (RI на рис. 11). Активи- рованные цАМФ-зависимые протеинкиназы изменяют метаболизм и функцио- нальную активность клеток, например в миокарде (бета1-адренорецепторы), — способствуют активации кальциевых каналов, входу кальция через клеточные мембраны и усилению всех функций сердца; в гладких мышцах (бета2-адреноре- цепторы) — способствуют расслаблению мышц, возможно, за счет фосфорилиро- вания (перевода в неактивную форму) киназы легких цепей миозина и облегчения удаления свободного кальция из цитоплазмы клетки во внутриклеточное депо. Дофаминовые рецепторы также сопряжены с аденилатциклазой. D1-рецепторы активируют аденилатциклазу и увеличивают содержание цАМФ в клетках; D2-рецепторы ингибируют аденилатциклазу. Роль дофамина и дофаминовых ре- цепторов в эфферентной иннервации не столь значительна, как в центральной нервной системе. Показано, что в некоторых подкорковых ядрах концентрация дофамина в 10 и более раз выше, чем норадреналина. Здесь выделяют по несколь- ко подтипов D1- и D2-рецепторов, имеющих свою локализацию и разное функ- циональное значение. Препараты, действующие на центральные дофаминовые и адренорецепторы, будут обсуждаться в других разделах учебника. Лекарственные средства, влияющие на адренергическую передачу, в зависи- мости от направленности своего действия разделяются на адреномиметики, адре- ноблокаторы (адренолитики) и симпатолитики. Препараты также подразделяют- ся по избирательности (селективности) действия на тот или иной тип и подтип адренорецепторов — это деление является чрезвычайно важным для практиче- ской медицины. Поэтому к изучению довольно громоздкой классификации адре- норецепторов с их особенностями локализации и выполняемых функций никак нельзя относиться поверхностно. АДРЕНОМИМЕТИКИ Вещества этой группы активируют адренорецепторы. Они различаются по из- бирательности действия на альфа- и бета-адренорецепторы и их подтипы, а так- же по силе и продолжительности действия. Адреномиметики могут возбуждать адренорецепторы непосредственно (препараты прямого действия) или действо- вать опосредованно через эндогенные катехоламины (препараты непрямого дей- ствия). К последним относятся препараты, которые способны освобождать мо- бильные запасы медиатора из нервных окончаний в синаптическую щель. Типич- ным представителем адреномиметиков непрямого действия является эфедрин. В зависимости от преобладающего влияния на тот или иной тип рецепторов адреномиметики прямого действия могут быть разделены на препараты, которые возбуждают: только альфа-адренорецепторы; только бета-адренорецепторы; оба типа рецепторов (смешанного действия) (табл. 32). Таблица32 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||