Бродский А.К. Краткий курс общей экологии. Учебное пособие. Спб. Деан. 2000. 224 с
 Скачать 2.74 Mb. Скачать 2.74 Mb.
|

|
89 активно питающиеся животные с неизбирательным питанием (грунтоеды, усатые киты, отдельные виды рыб и др.). Активно питающихся животных разделяют на две большие группы — «жнецы» (собиратели) и «охотники». Первые используют пищу, имеющуюся в изобилии, а вторые специализированы на добывании более редкого корма. Жнецы — это не только растительноядные животные; к ним относятся и многие «мирные» зоофаги (планктоноядные рыбы, усатые киты, многие насекомоядные птицы и звери и др.), использующие легко добываемую пищу, которую не нужно долго искать. Группа охотников включает не только хищников. Сюда входят те фитофаги, которые вынуждены отыскивать пищу (например, питающиеся семенами и плодами). Жнецы и охотники не полностью изолированные группы, между ними существуют постепенные переходы. Однако такое разделение на группы позволяет отчетливо выявить многие экологические закономерности. Так, для охотников характерно сложное территориальное поведение, а жнецы редко защищают свою территорию, но среди них чаще встречаются виды, ведущие сложный общественный образ жизни. Для жнецов характерны две формы питания. При пастьбе животные уничтожают только часть имеющихся в данном месте кормов и постепенно перемещаются на новые участки. Выедание отличается тем, что корм уничтожается весь или почти весь, и только после этого животные мигрируют в другие места. Охотники по формам питания разделяются на три типа. Многие хищники («засадники») подкарауливают свою добычу и нападают на нее неожиданно, обычно из укрытия (богомолы, щуки, ястребы, кошки и пр.). Другие животные («искатели»), например насекомоядные птицы, проводят значительную часть времени в поисках добычи, но преследуют, схватывают и поедают ее довольно быстро, расходуя на это сравнительно мало энергии. Охотники третьего типа («преследователи») затрачивают немного времени на поиски добычи, но овладение ею требует от них значительных усилий (многие крупные хищники, соколы и т. п.). Конечно, некоторые животные могут использовать все три способа охоты в зависимости от ситуации, возможны также промежуточные варианты. Способ добычи ограничивает круг возможных источников пищи для хищника, однако эти факторы не приводят к строгой монофагии. Абсолютная монофагия свойственна некоторым насекомым-парализаторам. В этом 90 случае, в связи со сложностью инстинктивных операций, направленных на ^обездвиживание жертвы, необходима очень тонкая и точная подгонка поведения хищника к строению тела жертвы. Добычей лангедокского сфекса служат неповоротливые крупные бескрылые кузнечики эфиппигеры. Сфекс, схватив свою жертву за грудной щит, наносит ей два последовательных укола жалом, сначала снизу в грудь, чтобы поразить средний и задний нервные узлы, а затем в горло, чтобы поразить переднегрудной узел. Песчаная аммофила при парализации гусеницы последовательно жалит ее снизу в каждый сегмент, постепенно перехватывая ее челюстями и передвигаясь от головы к заднему концу. Ярко выраженная пищевая специализация обнаруживается у многих паразитических животных. Факторами, обеспечивающими и поддерживающими узкую олигофагию, могут быть особенности поведения паразита и его способность развиваться в строго определенных хозяевах. Адаптированность каждого вида паразита к своему хозяину проявляется, во-первых, в морфологических изменениях, характерных для паразитического образа жизни, во-вторых, в развитии физиологических реакций подавления иммунитета и, в-третьих, в синхронизации циклов развития паразита и хозяина. Пищевая специализация фитофагов обычно основана на их способности реагировать на определенные химические компоненты растений. Стимулами, привлекающими или отталкивающими животных, могут быть различные органические соединения, которые содержатся в растительном субстрате, но которые, как правило, не имеют питательной ценности. Чаще всего функцию этих стимулов выполняют гликозиды или терпены. Химические вещества, привлекающие животных, называются аттрактантами, а отталкивающие— репеллентами. Растение употребляется в пищу, если оно содержит необходимые пищевые аттрактанты и не имеет достаточно сильных репеллентов. Обогащенная аттрактантами пища сильнее привлекает фитофагов и поглощается ими в большем 91 количестве. Поэтому существует прямая связь между степенью привлекательности пищи и ее «оптимальностью», т. е. способностью поддерживать высокие темпы роста животных при минимальной смертности. «Неоптимальная» пища может обладать столь же высокой питательной ценностью, но она лишена аттрактантов или имеет сильные репелленты и поэтому поглощается в меньшем количестве. Понятно, что при питании «неоптимальной» пищей рост и развитие животных замедляется, а смертность возрастает. Когда пищевые аттрактанты широко распространены в мире растений и нет достаточно сильных репеллентов, мы имеем дело с полифагией. Пищевая специализация животных-олигофагов может быть связана с ограниченным распространением аттрактантов или, наоборот, с широким распространением и аттрактантов и репеллентов. Наконец, строгая монофагия обусловливается аттрактантами, которые свойственны только одной небольшой группе растений. Когда пищевые аттрактанты свойственны лишь ограниченному кругу растений, то возникает олигофагия. Так, у гусениц и бабочек-белянок специализация на питании крестоцветными несомненно связана с привлекающим действием гликозидов горчичного масла. Помимо узкой олигофагии выделяют также аномальную олигофагию, т. е. питание растениями нескольких видов неблизкородственных родов. Аномальным олигофагом является бабочка тутовый шелкопряд, гусеницы которой питаются белой шелковицей и маклюрой. Для гусениц этого вида известен ряд привлекающих веществ, которые по своему действию делятся на три группы: дистантные пищевые аттрактанты, контактные пищевые аттрактанты, вещества, вызывающие заглатывание пищи. Первая группа веществ привлекает гусениц на расстоянии и вызывает их передвижение по направлению к источнику корма. Контактные пищевые аттрактанты вступают в действие при непосредственном соприкосновении гусениц с кормом и способствуют отгрызанию кусочка листа. Для проглатывания его необходимо присутствие третьей группы веществ. Таким образом, аномальная олигофагия возникает в том случае, когда питание 92 стимулируется большой группой различных веществ', которые редко присутствуют вместе в одном и том же растении. Различия между этими тремя экологическими группами фитофагов носят не качественный, а скорее количественный характер (рис. 5.4). Монофаги и в меньшей степени олигофаги связаны с определенной и ограниченной группой кормовых растений. Потребление этих растений обеспечивает оптимальную скорость развития, плодовитость и прочие процессы жизнедеятельности. Поэтому в данном случае лишь количество пищи служит лимитирующим фактором. Иначе обстоит дело с полифагами. Здесь ограничивающим фактором становится качество пищи, т. е. содержание в ней определенных веществ, необходимых для осуществления различных процессов жизнедеятельности. В зависимости от стадии жизненного цикла полифаг оказывает предпочтение тем или иным ее видам. Так, самкам шотландской куропатки перед размножением 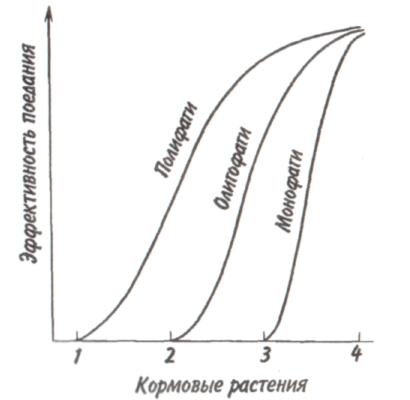 Рис. 5.4. Эффективность поедания кормовых растений тремя группами насекомых-фитофагов. Кормовые растения: 1 - отвергаемые, 2 - непривычные, 3 - замещающие, 4 - оптимальные. 93 необходимы такие элементы, как кальций, фосфор и азот. Поэтому птицы выбирают в качестве корма вереск, богатый этими элементами, не трогая растений, которые содержат их в недостаточном количестве. Чем уже спектр потребляемых организмом пищевых ресурсов, тем теснее должна быть пространственная связь этого организма со скоплениями его ресурсов или же тем больше должны быть затраты времени и энергии на поиски необходимых ресурсов. Наличие и доступность многих пищевых ресурсов зависят от времени года. Хорошим примером могут служить заросли дикой малины в лесах умеренного пояса. Зимой растения малины представляют собой скопление мелких сучьев, но весной на них во множестве появляются молодые, богатые белком почки, а затем листья. Зацветание влечет за собой кратковременное выделение нектара, а нектар — это совершенно новый ресурс, существующий, правда, лишь во время цветения. Со временем завязываются плоды, и плоды эти, созревающие и созревшие, представляют собой новый обильный источник корма. Все эти сезонные образования могут являться надежными ресурсами либо для растительных полифагов, которые в состоянии переключиться на другую пищу, когда «сезон малины» минует, либо для монофагов, у которых весь период активной жизни втиснут в соответствующий сезон и которые проводят всю остальную часть года в спячке, или диапаузе, не нуждаясь в пище. 94 Т е м а 6 БИОТИЧЕСКИЕ ФАКТОРЫ Под биотическими факторами понимают различные формы взаимодействия между особями и популяциями. Биотические факторы подразделяются на две группы: межвидовые и внутривидовые взаимодействия. 6.1. ГОМОТИПИЧЕСКИЕ РЕАКЦИИ Под гомотипическими реакциями понимают взаимодействия между особями одного вида. Жизнедеятельность животных и растений при совместном обитании в значительной степени зависит от численности и плотности популяции. В связи с этим большое экологическое значение имеют явления, связанные с так называемыми эффектами группы и массы. Эффект группы — это влияние группы как таковой и числа индивидуумов в группе на поведение, физиологию, развитие и размножение особей, вызванное восприятием присутствия особей своего вида через органы чувств. Многие насекомые (сверчки, тараканы, саранчовые и др.) в группе имеют более интенсивный, чем при жизни поодиночке, метаболизм, быстрее растут и созревают. Баклан —главный производитель гуано в Перу, может существовать лишь при условии, если в его колониях насчитывается не менее 10 000 особей и на 1 м2 приходится 3 гнезда. При совместной жизни легче искать и добывать пищу, а также защищаться от врагов. Объединенные в стаю волки способны убивать добычу более крупных размеров, чем действуя в одиночку. Бизоны, мускусные быки и другие жвачные успешнее обороняются от хищников, если они объединены в стада. Нередко эффект группы проявляется уже при совместном существовании двух животных. Он может приводить не только к положительным, но и к отрицательным 95 результатам. Например, рост головастиков в группе замедляется. Как правило, при небольшой численности группы преобладают положительные эффекты, а при избыточной плотности животных доминируют отрицательные явления. Так, в перенаселенных группах домовых мышей падает плодовитость и даже совсем прекращается размножение. У мучного хрущака при высокой плотности популяции намного увеличивается доля бесплодных яиц. Групповой эффект наиболее ярко выражается при наличии у животных явления фазности, т. е. способности вида существовать одновременно в двух формах — в форме одиночных особей и в форме групп. Эффект группы наглядно виден у мигрирующих саранчовых, которые при изолированном воспитании представляют собой одиночную фазу (солитария), окрашенную в зеленый цвет, а при скученном воспитании возникает стадная фаза (грегария), отличающаяся контрастными черными пятнами на оранжевом или светло-коричневом основном фоне. Эффект массы, в отличие от эффекта группы не связан с восприятием особями одного вида присутствия друг друга. Он выливается изменениями в среде обитания, происходящими при увеличении численности особей и плотности популяции. Как правило, эффект массы отрицательно сказывается на плодовитости, скорости роста, длительности жизни животных. Например, при развитии популяции мучного хрущака в муке постоянно накапливаются экскременты, линочные шкурки, что приводит к ухудшению муки как среды обитания. Это вызывает падение плодовитости и повышение смертности в популяции жуков. В природе эффекты группы и массы далеко не всегда легко различить, поскольку они нередко проявляются одновременно. Исключительно важную роль играют групповые и массовые эффекты в динамике численности популяций, выступая в ряду так называемых зависящих от плотности факторов, которые регулируют численность популяции по принципу обратной связи. 96 Здесь следует упомянуть принцип Олли: для каждого вида существует оптимальный размер группы и оптимальная плотность популяции. Как перенаселенность, так и недоселенность (или отсутствие агрегации) могут оказывать неблагоприятное влияние. Это положение подтверждено многочисленными наблюдениями и экспериментами. Известно, например, что для успешного выживания стадо слонов должно состоять по крайней мере из 25 особей, а стадо северного оленя — минимум из 300 голов. Гнездящиеся совместно чайки и другие птицы часто не могут успешно размножаться, если колония слишком мала. Однако и очень большая численность группы для них оказывается неблагоприятной. Таким образом, действие принципа Олли объясняется совместным влиянием эффектов группы и массы. К гомотипическим реакциям кроме группового и массового эффектов относится еще одна форма взаимодействия между особями одного вида — внутривидовая конкуренция. Сам термин появился сравнительно недавно. Раньше в том же значении использовался термин «борьба за существование», предложенный Ч. Дарвином, т. е. борьба за возможность выжить, для чего необходима энергия, получаемая растениями в виде солнечного света, а животными в виде различных пищевых материалов. Поэтому в борьбе за овладение источниками энергии происходит напряженная конкуренция, возникает соперничество между особями одного вида. В отношении всех видов конкуренции существует правило: чем более совпадают потребности конкурентов, тем острее конкуренция. Следовательно, внутривидовая конкуренция острее межвидовой, конкуренция между особями одной популяции острее, чем между особями разных популяций, и т.д. Победивший в конкурентной борьбе имеет возможность оставить потомство, а, следовательно, передать свои гены по наследству. Уступивший в конкурентной борьбе погибает или, точнее, элиминируется. Таким образом, основной результат, к которому приводит внутривидовая конкуренция,— дивергенция особей, завершающаяся формированием популяций (рис. 6.1). 97 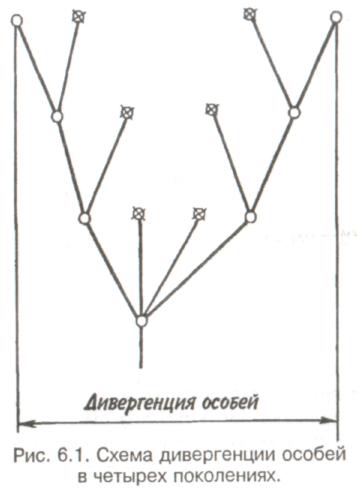 Перечеркнуты кружки, соответствующие особям, которые не оставили потомства. Различают две основные формы конкуренции — прямую и косвенную. Прямая конкуренция, или интерференция, осуществляется путем прямого влияния особей друг на друга, например, при агрессивных столкновениях между животными или при выделении токсинов (аллелопатия) у растений и микроорганизмов. Интерференция у животных не обязательно заключается во взаимной агрессивности. Она может возникать, например, когда одни животные своим поведением лишают других доступа к пище, укрытиям, к местам для размножения и т. д. Косвенная конкуренция не предполагает непосредственного взаимодействия между особями. Она происходит опосредованно — через потребление разными животными одного и того же ресурса, который обязательно должен быть ограниченным. Поэтому такую конкуренцию обычно называют эксплуатационной. В качестве причи- 98   ны, вызывающей конкуренцию, может выступать любой лимитированный ресурс, а также места для размножения, укрытия и т. д. При эксплуатационной конкуренции преимущество получают животные, способные быстрее и эффективнее использовать ресурсы, являющиеся предметом конкуренции, и уменьшать их количество, достающееся другим животным. ны, вызывающей конкуренцию, может выступать любой лимитированный ресурс, а также места для размножения, укрытия и т. д. При эксплуатационной конкуренции преимущество получают животные, способные быстрее и эффективнее использовать ресурсы, являющиеся предметом конкуренции, и уменьшать их количество, достающееся другим животным.Внутривидовая конкуренция нередко проявляется в территориальности—охране отдельными животными или их группами индивидуальных участков. Обычно охраняется кормовая территория. Это характерно для некоторых видов насекомых, рыб, многих видов ящериц, млекопитающих и подавляющего большинства видов птиц. Существуют также гнездовые и брачные территории, появление которых обусловлено наличием конкурентной борьбы за места размножения или за брачного партнера. Конкуренция за некоторый ограниченный ресурс обычно приводит к более равномерному пространственному размещению животных. У животных с общественным поведением конкуренция нередко выражается в возникновении иерархии доминирования, позволяющей упорядочить распределение ресурсов между животными в группе. Поскольку основным объектом внутривидовой конкуренции служит энергия, то среди растений наиболее острая борьба идет за свет. При слишком большой густоте произрастания растения затеняют друг друга. Конкуренция выражается также в гибели определенного количества растений, в результате чего плотность последних снижается. Это явление хорошо выражено у некоторых культур (морковь, свекла). Оно наблюдается и в лесу, где молодых деревьев гораздо больше, чем старых. Конкуренция за свет влияет на форму деревьев: у них совершенно различный вид в зависимости от того, растут ли они в лесу или поодиночке. Например, у дуба, если он растет отдельно, крона шарообразной формы, так как нижние боковые ветки получают достаточное количество света, чтобы развиваться не хуже верхних. В лесу же, напротив, листва нижних ветвей затенена вышерасположенными ветвями как этого дерева, так 99 и соседних растений. Происходит естественный процесс отмирания и опадения ветвей, листья которых получают мало света и у которых баланс ассимиляции и дыхания отрицательный. По мере роста в высоту опадение нижних ветвей происходит быстрее, и дерево принимает лесную форму, отличающуюся прямым цилиндрическим стволом, который на большом протяжении лишен ветвей. Конкуренция протекает, но не по принципу «включена-выключена». Напротив, ее уровень непрерывно меняется в зависимости от конкретных условий. Так, в экологическом вакууме конкуренция слабо выражена или отсутствует, а в насыщенной среде она играет большую роль во взаимоотношениях между особями. 6.2. ГЕТЕРОТИПИЧЕСКИЕ РЕАКЦИИ Гетеротипические реакции — это взаимодействия между особями, принадлежащими к разным видам. Две популяции либо влияют, либо не влияют друг на друга. Если влияние есть, то оно может быть благоприятным или неблагоприятным. Для рассмотрения гетеротипических реакции спекшим таблицу, где символами обозначены возможные взаимодействия. Различные типы взаимодействий между популяциями видов А и В 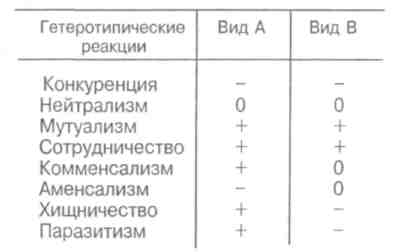 Примечание. «0» — нет влияния на данный вид, « + » — благоприятное влияние, «—» неблагоприятное (отрицательное) влияние. 100   Если каждая из двух популяций неблагоприятно влияет на другую, то взаимоотношения между ними носят характер конкуренции. В типичном случае двум популяциям требуется один и тот же ресурс (или ресурсы), имеющийся в недостатке. Если две популяции не взаимодействуют и ни одна из них не влияет на другую, то имеет место нейтрализм — случай, не представляющий особого экологического интереса. Истинный нейтрализм в природе очень редок, поскольку в любой экосистеме между всеми популяциями возможны косвенные взаимодействия. Приносящие обоюдную пользу взаимодействия относят к мутуализму, если объединение двух популяций облигатное (ни одна из сторон не может существовать без другой). В таком случае говорят о симбиозе, а популяции называют симбионтами. Если каждая из двух популяций неблагоприятно влияет на другую, то взаимоотношения между ними носят характер конкуренции. В типичном случае двум популяциям требуется один и тот же ресурс (или ресурсы), имеющийся в недостатке. Если две популяции не взаимодействуют и ни одна из них не влияет на другую, то имеет место нейтрализм — случай, не представляющий особого экологического интереса. Истинный нейтрализм в природе очень редок, поскольку в любой экосистеме между всеми популяциями возможны косвенные взаимодействия. Приносящие обоюдную пользу взаимодействия относят к мутуализму, если объединение двух популяций облигатное (ни одна из сторон не может существовать без другой). В таком случае говорят о симбиозе, а популяции называют симбионтами.Широко известным примером мутуализма могут служить лишайники, представляющие собой симбиоз водоросли и гриба. В животном мире пример наиболее совершенного симбиоза дают термиты, пищеварительный тракт которых служит приютом для жгутиковых. Термиты известны своей способностью питаться древесиной, несмотря на отсутствие у них ферментов, гидролизующих целлюлозу. Это делают за них жгутиконосцы. Образующийся сахар в основном используется простейшими, однако и термитам остается вполне достаточно. Термиты не могут существовать без этой кишечной фауны: свежевылупившийся термит инстинктивно слизывает жгутиконосцев с другой особи. При каждой линьке термиты лишаются всех жгутиконосцев, но, поскольку они живут колониями, каждая особь после линьки вновь получает их от соседа. В свою очередь жгутиконосцы снабжаются благодаря деятельности термитов необходимым количеством пищи и находятся в полной безопасности и в сравнительно постоянных условиях среды. Симбиоз может играть важную экологическую роль. Растения, живущие в симбиозе с бактериями — фиксаторами азота, обогащают почву азотистыми веществами. Древесные породы с фиксирующими азот клубеньками часто первыми заселяют бедные почвы. При сотрудничестве обе популяции образуют сообщество. Оно не является обязательным, так как каждый 101 вид может существовать изолированно, но жизнь в сообществе приносит обоим пользу. Иногда вместо термина «сотрудничество» употребляют термин «протокооперация». Совместное гнездование нескольких видов птиц, таких как крачки и цапли, представляет собой пример сотрудничества, позволяющего им более успешно защищаться от хищников. При совместном гнездовании белой цапли и розовой колпицы самцы цапель воруют ветки из гнезд колпиц. Подобные кражи иной раз приводят к дракам, но чаще робкие колпицы ограничиваются глухим горловым ворчанием — все же преимущества, благодаря такому образу жизни, перевешивают неприятности от посягательства цапель. Широко известен в литературе пример сотрудничества актинии с раком-отшельником. Актиния защищает рака-отшельника от врагов своими стрекательными щупальцами, а в ответ получает остатки его трапезы. В случае комменсализма, популяции одного вида сообщества (комменсал) извлекают пользу из сожительства, а популяции другого вида (хозяин) не имеют никакой выгоды. Отношения между комменсалами и хозяевами характеризуются взаимной терпимостью. Часто комменсал использует хозяина для передвижения. На наземных жуках, особенно на навозниках и мертвоедах, встречаются подвижные личинки гамазовых клещей, которые таким способом и расселяются. Кольчатый червь, живущий в раковине своего хозяина, рака-отшельника, питается остатками его пищи, схватывая их непосредственно с ротовых частей хозяина. Питание «остатками с хозяйского стола» может в конечном итоге привести к паразитизму, т. е. к питанию тканями хозяина. Об аменсализме говорят в тех случаях, когда одна популяция, именуемая аменсалом, претерпевает угнетение роста и размножения, а вторая, называемая ингибитором, таким испытаниям не подвержена. Аменсализм известен только у растений, грибов и бактерий. Благодаря выделению корнями токсических веществ ястребинка из семейства сложноцветных вытесняет другие однолет- 102 ние растения и образует чистые заросли на довольно больших площадях. Многие грибы и бактерии синтезируют антибиотики, тормозящие рост других бактерий. Хищничеством называют такое взаимодействие между популяциями, при котором одна из них, неблагоприятно влияя на другую, получает выгоду от этого взаимодействия. Обычно жертва убивается хищником и съедается целиком или частично (исключением служат ящерицы, оставляющие хищнику хвост, и растения, у которых травоядные объедают только листья). Паразитизм является, по существу, хищничеством, но хозяин (член популяции, испытывающей вредное влияние), как правило, не погибает сразу, а некоторое время используется паразитом. Следовательно, паразитизм можно рассматривать как ослабленную форму хищничества. Кроме того, между паразитизмом и хищничеством существуют переходные формы, которые трудно отнести к той или иной категории гетеротипических реакций. Например, среди насекомых есть виды-паразитоиды, которые вначале ведут себя как паразиты, щадя жизненно важные органы хозяина, а под конец развития, съедая своего хозяина, становятся настоящими хищниками. 6.3. ПРИНЦИП КОНКУРЕНТНОГО ИСКЛЮЧЕНИЯ. ЭКОЛОГИЧЕСКАЯ ДИВЕРСИФИКАЦИЯ Решающее значение в природе имеет межвидовая конкуренция, поскольку она в большей степени, чем другие гетеротипические реакции, определяет роль видов в экосистемах. Межвидовая конкуренция — это такое взаимодействие, когда два вида соперничают из-за одних и тех же источников существования — пищи, пространства и т. п. Причем она возникает в тех случаях, если использование источника ресурсов одним видом ведет к ограничению использования его другим. Внутривидовая конкуренция сильнее межвидовой, но правило конкуренции распространяется и на последнюю: конкуренция между двумя видами тем сильнее, чем бли- 103 же их потребности. В крайнем случае, можно допустить, что два вида с совершенно одинаковыми потребностями не могут существовать вместе: один из них через какое-то время обязательно будет вытеснен. Это положение получило статус закона, известного под названием принципа конкурентного вытеснения, или принципа Гаузе — в честь Г. Ф. Гаузе, впервые продемонстрировавшего это явление в экспериментах с инфузориями (рис. 6.2). При изолированном содержании численность популяций обоих видов инфузорий (рис. 6.2, кривые 1) 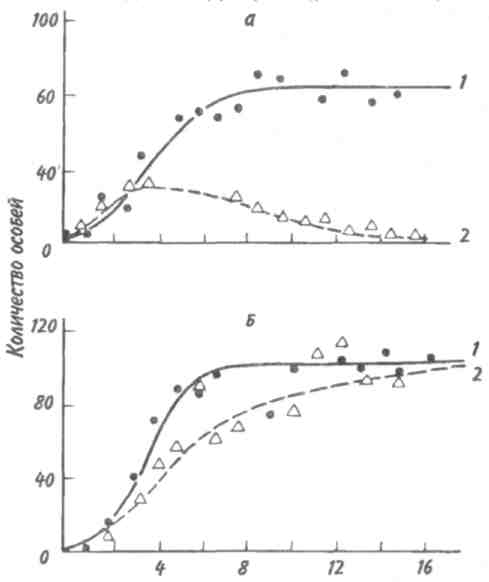 Рис. 6.2. Конкурентное исключение, продемонстрированное в лабораторном эксперименте на двух видах инфузорий (по Гаузе, 1934). a —Paramecium caudatum, б —P. aurelia: 1 — в изолированной культуре, 2—в смешанной культуре. 104 увеличивается до тех пор, пока не достигнет максимума. В смешанной культуре популяция второго вида (рис. 6.2,б, кривая 2) растет быстрее, чем первого (рис. 6.2,а, кривая 2), что приводит к ограничению ее численности. Если не произойдет изменений в. параметрах конкуренции, то конкурентное исключение (вымирание первой популяции)—лишь вопрос времени. Иногда два близких вида, имеющих одинаковые пищевые потребности, живут на одной территории, не конкурируя друг с другом. Такие, казалось бы, исключения из принципа Гаузе можно обнаружить среди птиц. В Англии большой баклан и хохлатый баклан совместно гнездятся на одних и тех же скалах и кормятся в одних и тех же водах, но они вылавливают разную пищу. Большой баклан ныряет глубоко и питается преимущественно бентическими животными (камбала, креветки), тогда как хохлатый баклан охотится в поверхностных водах на сельдевых рыб. Подобные факты, а их много, подводят нас к понятию экологической ниши, предложенному Элтоном еще в 1927 г. По образному выражению Одума, местообитание — это адрес вида, тогда как экологическая ниша — это род его занятий. Иными словами, знание экологической ниши дает возможность ответить на вопросы: где, чем и как питается вид, чьей добычей он сам является, каким образом и где он размножается и т. д. Современное определение звучит следующим образом: экологическая ниша — это роль вида в переносе энергии в экосистеме. Имеющиеся в экологической литературе трактовки данного понятия чрезвычайно разнообразны. Все же можно дать следующую наиболее общую формулировку: экологическая ниша — это общая сумма всех требований организма к условиям существования, включая занимаемое им пространство, функциональную роль в сообществе (например, трофический статус) и его толерантность по отношению к факторам среды — температуре, влажности, кислотности, составу почвы и др. 105 Названные три критерия определения экологической ниши удобно обозначать как пространственную, трофическую и многомерную ниши. Пространственная ниша, или ниша местообитания, может быть названа «адресом» организма. Трофическая ниша характеризует особенности питания и, следовательно, роль организма в сообществе, как бы его «профессию». Многомерная, или гиперпространственная, ниша — это диапазон всех условий, при которых живет и воспроизводит себя особь или популяция. Для интерпретации этого понятия используется теория множеств. Многомерная ниша — это область в гиперпространстве, измерениями которого служат различные экологические факторы; она охватывает диапазоны толерантности по каждому фактору-измерению. Такой акцент исключает из рассмотрения функциональный и поведенческий аспекты ниши. Различают фундаментальную (потенциальную) нишу, которую организм мог бы занимать в отсутствие конкурентов, хищников и других врагов и в которой физические условия оптимальны, и реализованную нишу — фактический диапазон условий существования организма, который или меньше фундаментальной ниши, или равен ей. Фундаментальную нишу называют иногда преконкурентной, а реализованную — постконкурентной. Однако это не синонимы, поскольку не только конкуренция ограничивает ширину реализованной ниши. Взаимоотношение между фундаментальной и реализованной нишами может быть пояснено с помощью так называемой лицензионной модели. Понятие «экологической лицензии» было впервые введено Гюнтером, но оно было весьма расплывчатым и определялось как условия внешней и внутренней среды, разрешающие осуществляться некоторым эволюционным факторам и событиям. Позднее В. Ф. Левченко и Я. И. Старобогатов вернулись к понятию экологической лицензии, причем были учтены следующие особенности: место в пространстве и времени, роль потоков вещества и энергии и наличие градиентов внешних условий, обеспечиваемых экосистемой для популяций и организмов там существующих. 106 Приведенное описание лицензии напоминает те слова, которые используют, когда говорят о так называемой «пустой экологической нише». Однако, строго говоря, последняя не может быть объяснена, если использовать классические определения ниши. Любая популяция имеет фундаментальную (потенциально возможную) и реализованную (фактическую) экологическую нишу (рис. 6.3). Реализованная ниша RNникогда не выходит за границы лицензии L, но при этом обязательно перекрывает фундаментальную нишу FN. Если внутри лицензий экосистемы находится по одной популяции, то мы имеем дело со случаем простой экоси- 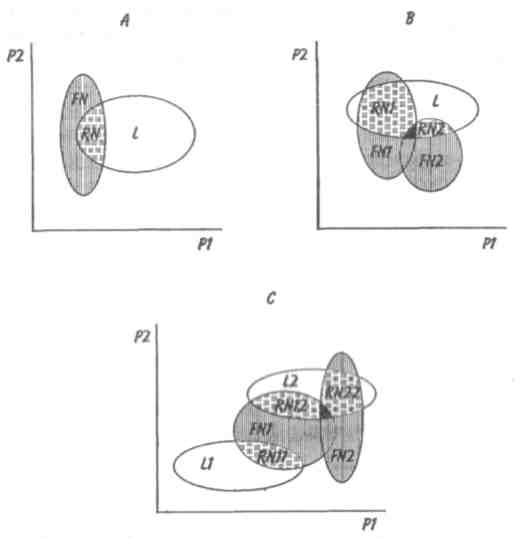 Рис. 6.3. Взаимодействия между фундаментальными нишами Видов (FN), реализованными нишами популяций (RN) и лицензиями (L.) в пространстве внешних факторов (Р1 и Р2). А-С — различные варианты взаимодействия ниш и лицензий. Реализованные и фундаментальные ниши различным образом затенены, зачерненные фрагменты — области конкуренции (по Левченко, 1993). 107 стемы. При наличии в лицензии нескольких популяций между ними возможна конкуренция за ресурсы. В этом случае мы имеем случай сложной экосистемы. Для сложной экосистемы можно говорить о том, что у нее имеются групповые фундаментальные ниши, а в лицензии каждого трофического уровня существуют групповые реализованные ниши. Теперь конкретизируем принцип Гаузе: два вида не могут занимать одну и ту же экологическую нишу. Принцип Гаузе явился важным экологическим обобщением. Он позволил увидеть, что для сосуществования видов в конкурентных сообществах абсолютно необходимы какие-то различия их экологических ниш. Поскольку для каждой из конкурирующих сторон избегание взаимодействия выгодно, конкуренция играет роль мощного эволюционного фактора, приводящего к разделению ниш, специализации видов и возникновению видового разнообразия. Явление разделения экологических ниш в результате межвидовой конкуренции получило название экологической диверсификации. Экологическая диверсификация между обитающими совместно (т.е. симпатрическими) видами в большинстве случаев осуществляется по следующим трем параметрам: пространственному размещению, пищевому рациону и распределению активности во времени. Как правило, для значительного ослабления и даже полного устранения конкуренции достаточно возникновения четких различий по одному из этих параметров. Различия в пространственном размещении заключаются в выборе видами разных стаций или даже микростаций. Большой пестрый дятел ищет пищу в основном на стволах деревьев, средний пестрый — на больших ветвях, а малый пестрый дятел — на ветках кроны. Благодаря пути из варяг в греки на север проникла черная крыса. Она загнала серую крысу в подполье, а сама обосновалась на чердаках. Когда в связи с развитием судоходства на юге появилась серая крыса, или пасюк, то она вытеснила черную крысу на чердачные помещения, а сама обосновалась в подполье и канализационной сети. 108 Экологическая диверсификация по пищевому рациону— широко распространенное явление. Не раз было показано, что близкородственные виды животных характеризуются различными пищевыми потребностями. Пищевые рационы пустынных ящериц, принадлежащих к нескольким симпатрическим видам, состоят у одних преимущественно из муравьев, у других из термитов, у третьих из ящериц других видов или только растений. Сходные примеры различий в рационах у родственных симпатрических видов известны для многих птиц и млекопитающих. Разделение ниш часто бывает сопряжено с распределением активности видов во времени. Примерами могут служить разные типы суточной (ласточки и летучие мыши) или сезонной (некоторые виды ящериц, насекомых) активности. В умеренных широтах у садовой и черноголовой славок наблюдается сдвиг в периодах кладки в две недели. В тропиках эти виды могут размножаться круглый год, так как постоянное обилие необходимых для их питания насекомых ослабляет конкуренцию. Конкуренция за пищу приводит иногда к ускорению развития и изменению всего жизненного цикла вида. Интересный пример такой конкуренции представляют муха-саркофагида и оса-сфекс. Саркофагида заражает саранчовое, погружая яйцо в его тело. Сфекс парализует ослабленное саранчовое, после чего затаскивает его, а заодно и яйцо саркофагиды, в свою норку/ Обычно личинка паразитической мухи некоторое время развивается, но затем погибает, не выдержав пищевой конкуренции с быстро растущей личинкой сфекса. Однако отдельные личинки саркофагид благодаря ускорению развития успевали съесть парализованное саранчовое до появления личинки осы. Поскольку личинки саркофагид попадают в норку сфекса более или менее регулярно, то в эволюции закрепилась более высокая скорость развития личинок саркофагид, следствием чего стала меньшая плодовитость мух. Одним из следствий экологической диверсификации является так называемое смещение признаков. Иногда особи двух широко распространенных видов более сходны в тех частях своих ареалов, в которых встречаются 109 по отдельности (аллопатрически), чем на участках совместного обитания. Например, два вида поползней столь похожи друг на друга в районах аллопатрии, что различить их может только специалист. Но в областях совместного обитания они имеют специфические признаки: у одного клюв и черная заглазничная полоса больше, чем у другого. Исторически возникшее приспособление к наиболее полному использованию условий местообитания и к пространственному расселению у животных и растений привело к образованию жизненных форм. Их группирование по тем или иным признакам являет собой примеры экологической классификации растительного и животного мира. Если систематическая классификация растений и животных строится на основе родства организмов, то экологическая классификация (т. е. классификация жизненных форм) выявляет возможные пути приспособления организмов к среде обитания. В качестве примера жизненных форм у растений можно привести травы, кустарнички, кустарники, деревья и т. д., а среди животных — плавающие формы, роющие формы и т. п. Жизненные формы организмов следует отличать от экологических групп организмов. Если первые отражают приспособленность организмов к специфике данного местообитания в целом, то экологические группы отражают приспособленность организмов к отдельным факторам внешней среды. Например, по отношению к условиям увлажнения выделяются экогруппы — мезофиты, ксерофиты, суккуленты и т. д., к условиям освещения — светолюбы и тенелюбы, по отношению к типам почв — криофиты (жители холодных почв), галофиты (солерсы) и т. д. Представители одной и той же жизненной формы могут принадлежать к разным экологическим группам. Например, клевер луговой и астрагал выделяющийся относятся к стержнекорневым растениям, но принадлежат к разным экологическим группам: клевер, произрастающий на лугах, является мезофитом, астрагал, обитающий в сухостепных районах,— ксерофитом. Манжетка обыкновенная и копытень европейский, будучи представителями короткокорневищных растений, по отношению к свету распределяются по разным экогруппам: манжетка является светолюбом, а копытень — тенелюбом. В то же время оба 110 вида по отношению к фактору влажности принадлежат к одной экологической группе — мезофитам. На успехе в конкурентной борьбе того или иного вида сказывается влияние различных абиотических факторов. Воздействие среды на исход конкуренции изучалось в многочисленных лабораторных экспериментах с двумя видами малого мучного хрущака, Triboliumcastaneumи T.confuswn. Было установлено, что при равенстве исходной численности жуков обоих видов в жарком влажном климате (t0 = 34°С, относительная влажность 70%) всегда побеждает Т.castaneum. При ином соотношении начальной численности популяций независимо от факторов среды успех сопутствует то одному, то другому виду (рис. 6.4). Первоначальное доминирование какого-либо вида увеличивает вероятность его победы. При незначительном перевесе в численности той или иной популяции (переходная зона) каждый из видов может одержать победу в зависимости от условий температуры и влажности. 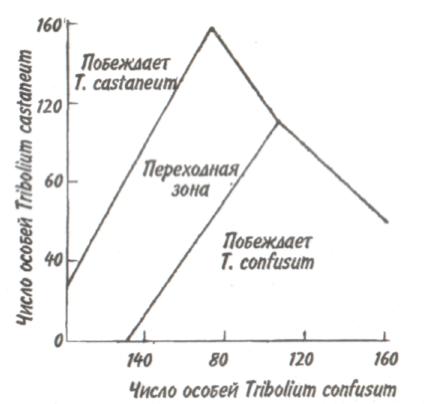 Рис. 6.4. Зависимость исхода конкуренции между лабораторными линиями жуков TriboliumcastaneumИ Т. confusumот начальных плотностей особей (из Пианка, 1981). 111 Оценивая успех в конкурентной борьбе одного из двух видов в природных условиях, опираться на такой показатель, как исходная плотность популяции, не представляется возможным. Как правило, вид более специализированный, стенобионтный, оказывается и более конкурентоспособным по сравнению с видом эврибионтным. Один из двух видов плоских червей обитает только в верховьях рек при температуре воды от 6,5 до 16 °С, а другой вид, выносящий температуры в пределах от 6,5 до 23 °С, может жить на всем протяжении реки. Когда оба вида симпатрические, то стенотермный вид вытесняет эвритермный из верховьев, и последний сохраняется только в нижнем течении реки, где значения температуры воды колеблются между 13 и 23 °С. На Кубе обитают два вида крокодилов: острорылый и кубинский. Несмотря на общие биологические особенности они, за редким исключением, никогда вместе не встречаются. Острорылый крокодил имеет большую экологическую пластичность, он хорошо переносит осолонение водоемов, поэтому широко распространен во многих прибрежных водоемах: устьях рек, лагунах и других низких местах. В отличие от острорылого кубинский крокодил встречается только в пресных водах. При совместном обитании он как более специализированный и поэтому более конкурентоспособный вытесняет острорылого крокодила из пресных водоемов, принуждая его переходить в солоноватые воды. Важным параметром вида является ширина его экологической ниши. Этот показатель всегда относителен. Мы можем говорить о широкой или узкой нише какого-нибудь организма лишь в сравнении с другими. Например, гусеницы дубовой листовертки, питающиеся только листьями дуба, имеют более узкую пищевую нишу, чем гусеницы непарного шелкопряда, развивающиеся на многих видах лиственных деревьев. Эврибионтные организмы обычно имеют более широкие ниши, чем стенобионтные. Однако экологическая ниша может иметь различную ширину по разным измерениям, таким как пространственное распределение, трофические связи и др. 112 Когда два организма различных видов используют одни и те же ресурсы, их ниши перекрываются. Перекрывание может быть полным или частичным, может быть по одному измерению ниши, но не быть по другим. Если какие-то ресурсы имеются в недостаточном количестве, то при перекрывании ниш возникает конкуренция, которая тем интенсивнее, чем сильнее это перекрывание. На рис. 6.5 показаны разные варианты взаимодействия экологических ниш. В случае, когда ниши полностью разобщены (рис. 6.5, в), населяющие их виды не конкурируют друг с другом. Если они вообще не вступают в конкуренцию с другими организмами, то их ниши близки к потенциально возможным в данной ситуации. Когда экологические ниши частично совпадают (рис. 6.5,6), сосуществование видов становится возможным благодаря специфичности их приспособлений, 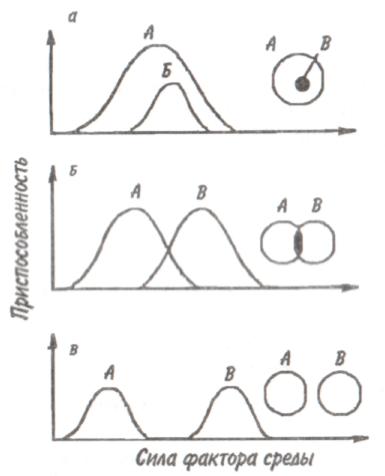 Рис. 6.5. Примеры возможных взаимодействий экологических ниш видов А и В (по: Пианка, 1981). Объяснения в тексте. 113 например, особи одного вида занимают убежища, недоступные для более активного конкурента. Наконец, когда ниша одного вида включает в себя нишу второго вида или эти ниши полностью перекрываются (рис. 6.5,а), произойдет конкурентное исключение одного из видов или доминирующий конкурент оттеснит своего соперника на периферию зоны приспособленности. Реальная экологическая ниша слабого конкурента предельно сузится, заняв лишь некоторую часть потенциально возможной ниши. Именно поэтому целый ряд животных и растений живет в условиях, отличных от оптимальных. Например, многие растения, принимаемые за кальциефобные, переносят в чистой культуре самое различное содержание кальция; в природных условиях эти растения сдерживаются в границах своего ареала конкуренцией других видов. Конкуренция приводит к очень важным экологическим последствиям. В природе особи каждого вида являются объектами одновременно межвидовой и внутривидовой конкуренции. Межвидовая конкуренция оказывает прямо противоположное, по сравнению с внутривидовой, влияние на пределы выносливости популяции, способствуя сужению диапазона используемых ею местообитаний и ресурсов. Это происходит потому, что разные виды обычно имеют различную способность осваивать 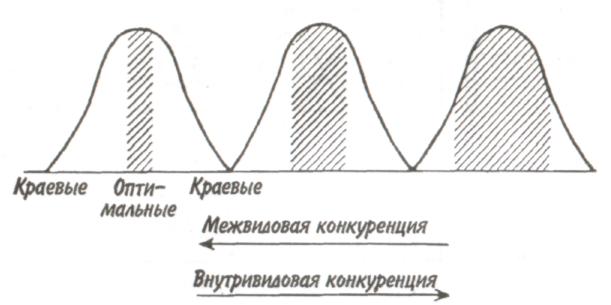 Рис. 6.6. Влияние межвидовой и внутривидовой конкуренции на распределение местообитаний у птиц (из Одума, 1975). 114 местообитания и потреблять ресурсы. При усилении внутривидовой конкуренции происходит дифференциация вида; он занимает большую территорию, распространяясь на менее благоприятные участки ареала (рис. 6.6). Расширяется экологическая ниша. Если преобладает межвидовая конкуренция, то происходит обратное: ареал уменьшается до территории с оптимальными условиями; одновременно увеличивается специализация (однообразие) вида. 115 |