Бродский А.К. Краткий курс общей экологии. Учебное пособие. Спб. Деан. 2000. 224 с
 Скачать 2.74 Mb. Скачать 2.74 Mb.
|

|
Некоторые характерные признаки г- и /К-отбора (Пианка, 1981)  128 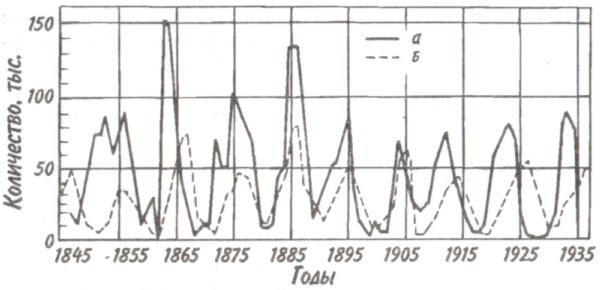  Рис. 7.9. Периодические колебания Рис. 7.9. Периодические колебанияпопуляций зайца-беляка (а) и рыси (б), установленные по числу шкурок, заготовленных «Компанией Гудзонова залива» (из Дажо, 1975). сравнению с численностью рыси обычно сдвинут на 1 -2 года назад. Это вполне понятно: рысь питается зайцами, а потому колебания ее численности должны быть связаны с колебанием численности ее добычи. Циклические изменения численности со средним периодом в 4 года характерны для обитателей тундры: полярной совы, песца, а также лемминга. По мнению многих ученых, периодичность 9,6-летних циклов у зайца-беляка и рыси определяется явлениями, происходящими в космосе, и так или иначе связана с солнечными циклами. Подобная зависимость отмечается, например, у атлантического канадского лосося, максимум численности которого наблюдается через каждые 9-10 лет. Причины, вызывающие другие периодические флуктуации численности, хорошо известны. У берегов Перу наблюдается трансгрессия теплых вод к югу. известная под названием Nino. Приблизительно один раз в 7 лет теплые воды вытесняют с поверхности холодные. Температура воды быстро поднимается на 5°С, изменяется соленость, гибнет планктон, насыщая воду продуктами распада. В результате погибает рыба, а за ней морские птицы. Случаи сезонных изменений численности популяций хорошо известны всем. Тучи комаров, большое 129 количество населяющих леса птиц обычно наблюдаются в определенный период года. В другие сезоны популяции этих видов могут практически исчезать. 7.4. ФАКТОРЫ ДИНАМИКИ ЧИСЛЕННОСТИ ПОПУЛЯЦИЙ Известно три типа зависимости численности популяции от ее плотности (рис. 7.10). При первом типе (кривая 1) скорость роста популяции уменьшается по мере увеличения плотности. Это широко распространенное явление позволяет понять, почему популяции некоторых животных относительно устойчивы. Во-первых, при увеличении плотности популяции наблюдается снижение рождаемости. Так, в популяции большой синицы при плотности меньше одной пары на 1 га на одно гнездо приходится 14 птенцов; когда же плотность достигает 18 пар на 1 га, выводок составляет менее 8 птенцов. Во-вторых, при увеличении плотности популяции меняется возраст наступления половой зрелости. Например, африканский слон в зависимости от плотности популяции может достигать половой зрелости в возрасте от 12 до 18 лет. Кроме того, этот вид при низкой плотности дает приплод 1 слоненок за 4 года, тогда как при высокой — рождаемость составляет 1 слоненок за 7 лет. При втором типе зависимости (кривая 2) темп роста популяции максимален при средних, а не при низких 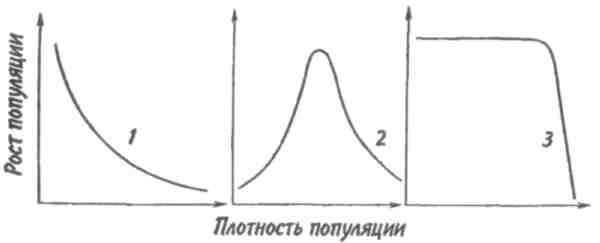 Рис. 7.10. Три типа зависимости роста популяции от плотности. Объяснение в тексте. 130  значениях плотности. Так, у некоторых видов птиц (например, чаек) число птенцов в выводке увеличивается с повышением плотности популяции, а затем, достигнув наибольшей величины, начинает уменьшаться. Этот тип влияния плотности популяции на скорость размножения особей характерен для видов, у которых отмечен групповой эффект. При третьем типе (кривая 3) темп роста популяции не изменяется до тех пор, пока она не достигнет высокой плотности, затем резко падает. значениях плотности. Так, у некоторых видов птиц (например, чаек) число птенцов в выводке увеличивается с повышением плотности популяции, а затем, достигнув наибольшей величины, начинает уменьшаться. Этот тип влияния плотности популяции на скорость размножения особей характерен для видов, у которых отмечен групповой эффект. При третьем типе (кривая 3) темп роста популяции не изменяется до тех пор, пока она не достигнет высокой плотности, затем резко падает.Подобная картина наблюдается, например, у леммингов. При пике численности плотность леммингов становится избыточной, и они начинают мигрировать. Элтон так описал миграции леммингов в Норвегии: животные проходили через деревни в таком количестве, что собаки и кошки, которые вначале нападали на них, просто перестали их замечать. Достигнув моря, обессиленные лемминги тонули. Регуляция численности равновесных популяций определяется преимущественно биотическими факторами. Среди них главным фактором часто оказывается внутривидовая конкуренция. Примером может служить борьба птиц за места для гнездования. Внутривидовая конкуренция может быть причиной физиологического эффекта, известного под названием шоковой болезни. Его отмечают, в частности, у грызунов. Когда плотность популяции становится слишком большой, шоковая болезнь приводит к снижению плодовитости и увеличению смертности, что возвращает плотность популяции к нормальному уровню. У некоторых видов животных взрослые особи питаются собственным приплодом. Это явление, известное как каннибализм, снижает численность популяции. Каннибализм свойствен, например, окуням: в озерах Западной Сибири 80% пищи крупных особей составляет молодь того же вида. Молодь, в свою очередь, питается планктоном. Таким образом, когда нет других видов рыб, взрослые особи живут за счет планктона. Межвидовые взаимодействия также играют существенную роль в контроле плотности популяции. Взаимодействия паразит-хозяин и хищник-жертва часто за- 131 висят от плотности. Болезни также являются фактором, принимающим участие в регуляции плотности популяции. Когда кролики болеют миксоматозом, вызываемым вирусом, распространение инфекции идет гораздо быстрее в популяциях с повышенной плотностью. Хищничество как ограничивающий фактор само по себе имеет большое значение. Причем если влияние жертвы на численность популяции хищника не вызывает сомнений, то обратное воздействие, т. е. на популяцию жертвы, бывает не всегда. Во-первых, хищник уничтожает больных животных, тем самым он улучшает средний качественный состав популяции жертвы. Во-вторых, роль хищника ощутима только тогда, когда оба вида обладают приблизительно одинаковым биотическим потенциалом. В противном случае из-за низкого темпа размножения хищник не в состоянии ограничить численность своей жертвы. Например, только одни насекомоядные птицы не могут остановить массовое размножение насекомых. Иными словами, если биотический потенциал хищника намного ниже биотического потенциала жертвы, действие хищника приобретает постоянный характер, не зависящий от плотности его популяции. Численность насекомых-фитофагов нередко определяется комбинацией видоспецифичных реакций насекомых и растений на воздействие загрязняющих веществ. Загрязнение понижает резистентность растений, в результате чего численность насекомых возрастает. Однако при слишком большом загрязнении численность насекомых падает, несмотря на уменьшение сопротивляемости растений. Приведенная дифференциация факторов динамики численности популяций позволяет понять их реальное значение в жизни и воспроизводстве популяций. Современная концепция автоматического регулирования численности популяций базируется на сочетании двух принципиально различных явлений: модификаций, или случайных колебаний численности, и регуляций, действующих по принципу кибернетической обратной связи и нивелирующих колебания. В соответствии с этим выделяют модифицирующие (не зависящие от плотности популяции) и регулирующие (зависящие от плотности по- 132 пуляции) экологические факторы, причем первые из них воздействуют на организмы либо непосредственно, либо через изменения других компонентов биоценоза. По существу, модифицирующие факторы представляют собой различные абиотические факторы. Регулирующие факторы связаны с существованием и активностью живых организмов (биотические факторы), поскольку лишь живые существа способны реагировать на плотность своей популяции и популяций других видов по принципу отрицательной обратной связи (рис. 7.11). 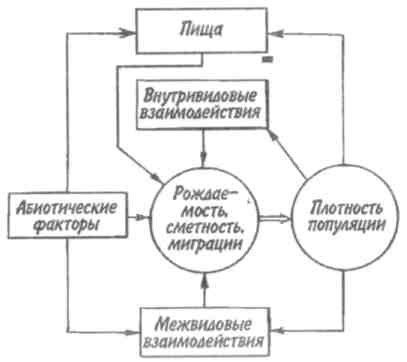 Рис. 7.11. Факторы динамики численности популяции с позиций концепции автоматического регулирования (по Викторову, 1976). Если воздействия модифицирующих факторов приводят лишь к преобразованиям (модификациям) колебаний численности, не устраняя их, то регулирующие факторы, выравнивая случайные отклонения, стабилизируют (регулируют) численность на определенном уровне. Однако на разных уровнях численности популяции регулирующие факторы принципиально различны (рис. 7.12). Например, хищники-полифаги, способные при изменении численности жертв ослабить или усилить свою активность — функциональнаяреакция—оказывают дей- 133 ствие при сравнительно низких значениях численности популяции жертвы. Хищники-олигофаги, характеризующиеся в отличие от полифагов численной реакцией на состояние популяции жертвы, оказывают на нее регулирующее действие в более широком диапазоне, чем полифаги. При достижении популяцией жертвы еще более высокой численности создаются условия для распространения болезней и, наконец, предельный фактор регуляции —внутривидовая конкуренция, ведущая к исчерпыванию доступных ресурсов и развитию стрессовых реакций в популяции жертвы. На рис. 7.12 представлена многозвенная буферная система регулирования численности популяции под воздействием биотических факторов, степень влияния которых зависит от плотности популяции. В реальной ситуации данный параметр зависит от большого числа факторов, в частности и тех, которые не оказывают регулирующего воздействия на плотность популяции по принципу обратной связи. Взаимодействие между модифицирующими, регулирующими, а также такими специфическими факторами, как размеры тела, группы и индивидуального уча- 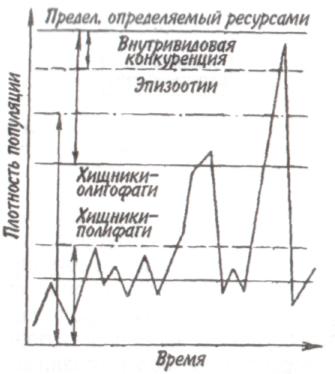 Рис. 7.12. Включение разных регулирующих механизмов на разных уровнях численности популяции (по Викторову, 1976). 134  стка, при их влиянии на плотность популяции млекопитающих показано на рис. 7.13. стка, при их влиянии на плотность популяции млекопитающих показано на рис. 7.13.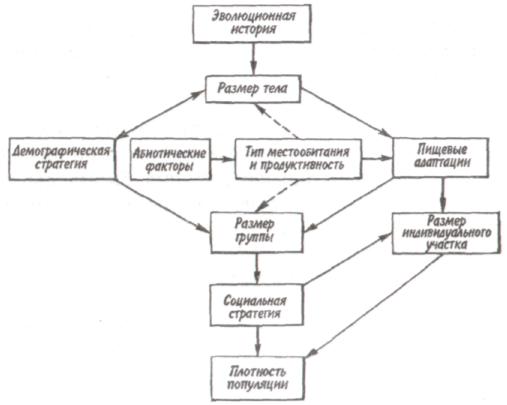 Рис. 7.13. Влияние размеров тела, факторов среды и поведения на плотность гипотетической популяции млекопитающих (по Эйзенбергу, 1983). Таким образом, чтобы получить исчерпывающую информацию о том, какие факторы вызывают колебания численности, теоретически требуются данные о физико-химических условиях, обеспеченности ресурсами, жизненном цикле этих организмов и влиянии конкурентов, хищников, паразитов и т. д., причем нужно знать, как все эти факторы влияют на рождаемость, смертность и миграцию. Все популяции непрерывно изменяются: новые организмы рождаются или прибывают как иммигранты, а прежние гибнут или эмигрируют. Несмотря на это, флуктуации размера популяции не безграничны. С одной стороны, она не может расти беспредельно, а с другой — вымирают виды достаточно редко. Следовательно, один из основных признаков популяционной динамики — сочетание изменений с относительной стабильностью. При этом колебания размеров популяций сильно различаются у разных видов. 135 Т е м а 8 ЭКОЛОГИЧЕСКАЯ СИСТЕМА На любом участке земной поверхности обитает всегда комплекс видов. В изоляции вид быстро ухудшает условия своего существования, поскольку увеличение биомассы происходит до тех пор, пока пищевые ресурсы не исчерпаны. После этого начнется отмирание биомассы. Если же дать дополнительные пищевые ресурсы, то и в таком случае падения биомассы избежать не удастся, так как будут накапливаться продукты обмена. 8.1. КОНЦЕПЦИЯ ЭКОСИСТЕМЫ Первые организмы на Земле были гетеротрофами. Они быстро исчерпали бы себя, если бы не появились автотрофы. При наличии этих групп организмов уже возможен примитивный круговорот веществ: Автотрофы синтезируют органические вещества, а гетеротрофы их потребляют. При этом происходит расщепление органических веществ. Если продукты расщепления вновь используются автотрофами, возникает круговорот между организмами, населяющими экосистему. Биотическую и абиотическую части экосистемы связывает непрерывный обмен материалом — круговороты питательных веществ, энергию для которых поставляет Солнце (рис. 8.1). Растения синтезируют органические соединения, используя энергию солнечного света и питательные вещества из почвы и воды. Эти соединения служат растениям строительным материалом, из которого они образуют свои ткани, и источником энергии, необходимой им для поддержания своих функций. Для высвобождения запасенной ими химической энергии гетеротрофы разлагают органические соединения на исходные неор- 136 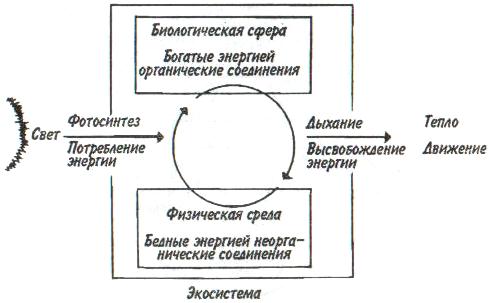 Рис 8.1. Поток энергии и круговорот химических веществ в экосистеме (по Риклефсу, 1979). ганические компоненты - диоксид углерода, воду, нитраты, фосфаты и т. п., завершая тем самым круговорот питательных веществ. Сказанное выше позволяет нам определить экосистему так: экологическаясистемапредставляет собой любое непрерывно меняющееся единство, включающее все организмы на данном участке и взаимодействующее с физической средой таким образом, что поток энергии создает определенную трофическую структуру, видовое разнообразие и круговорот веществ внутри системы. Другая формулировка звучит следующим образом: экосистема - исторически сложившаяся система совместного использования совокупностью живых организмов определенного пространства обитания в целях питания, роста и размножения. Экосистема есть основная функциональная единица живой природы, включающая и организмы, и абиотическую среду, причем каждая из частей влияет на другую и обе необходимы для поддержания жизни в том виде, в каком она существует на Земле. Двуединый характер этого комплекса подчеркнул В.Н. Сукачев в учении о биогеоценозе. Идеи, развиваемые Сукачевым, нашли графическое выражение на рис. 8.2. 137 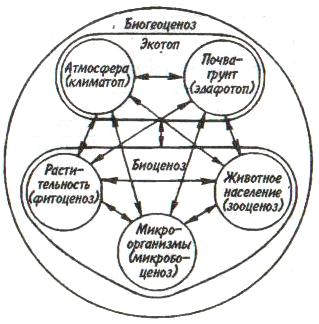 Рис. 8.2. Структура-биогеоценоза по Сукачеву (1964). Принимая двуединый характер биогеоценоза (экотоп + биоценоз), следует подчеркнуть, что неправомерно рассматривать биоценоз как сумму фитоценоза, зооценоза и микробоценоза, реально не существующих в природе в качестве отдельных и самостоятельных групп растений, животных и микроорганизмов. В современной экологической литературе экотоп часто обозначают как косную часть экосистемы, а биоценоз - как ее живую часть (рис. 8.3). 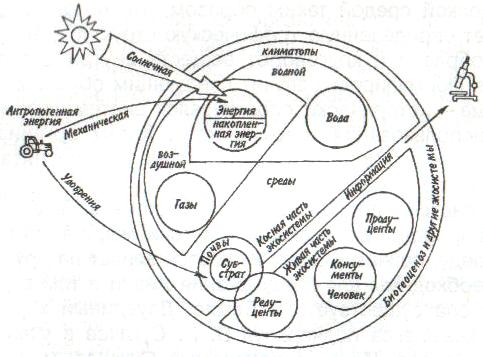 Рис. 8.З. Основные экологические компоненты биогеоценоза (из Реймерса, 1988). 138  В первом приближении биотическая часть экосистемы обязательно включает два основных компонента: В первом приближении биотическая часть экосистемы обязательно включает два основных компонента:
Функционирование автотрофов и гетеротрофов разделено также во времени: использование продукции ав-тотрофных организмов гетеротрофными может происходить не сразу, а с существенной задержкой. Например, в лесной экосистеме фотосинтез превалирует в листовом пологе. Лишь часть продуктов, причем весьма небольшая, немедленно и непосредственно перерабатывается гетеротрофами, питающимися листвой и молодой древесиной. Основная масса синтезированного вещества (в форме листьев, древесины и запасных питательных веществ в семенах, корнях) в конце концов, попадает в подстилку и почву, где и происходит утилизация органического вещества. С точки зрения их роли в экосистемах переходную группу между автотрофами и гетеротрофами образуют хемосин-тезирующие бактерии. Они получают энергию, необходимую для включения углекислого газа в состав компонентов клетки, не путем фотосинтеза, а в результате химического окисления таких простых неорганических соединений, как аммоний (окисляется в нитрит), нитрит (в нитрат), сульфид (в серу), закись железа (в оксид железа). Часть бактерий может развиваться в темноте, но большинство нуждается в кислороде. 139 Во втором приближении во всякой экосистеме можно выделить следующие компоненты: 1) неорганические вещества(углерод, азот, углекислый газ, вода и т. д.), вступающие в круговороты; 2) органическиесоединения (белки, углеводы, липиды, гуминовые вещества и т. д.), связывающие биотическую и абиотическую части; 3) климатическийрежим(температура и другие физические факторы); 4) продуценты—автотрофные организмы, главным образом зеленые растения, которые способны создавать пищу из простых неорганических веществ; 5) консументы—гетеротрофные организмы, главным образом животные, которые поедают другие организмы или частицы органического вещества; 6) редуценты (деструкторы, декомпозиторы) — гетеротрофные организмы, преимущественно бактерии и грибы, которые расщепляют сложные соединения до простых, пригодных для использования продуцентами. Первые три группы - неживые компоненты, а остальные составляют живой вес (биомассу). Расположение трех последних компонентов относительно потока поступающей энергии представляет собой структуру экосистемы (рис 8.4). Продуценты улавливают солнечную энергию и переводят ее в энергию химических связей. Консументы, поедая продуцентов, разрывают эти связи. Высвобожденная энергия используется консументами для построения собственного тела. Наконец, редуценты рвут химические связи разлагающегося органического вещества и строят свое тело. В результате вся энергия, запасенная продуцентами, оказывается использованной. Органические вещества разлагаются на неорганические и возвращаются к продуцентам. Таким образом, структуру экосистемы образуют три уровня (продуценты, консументы, редуценты) трансформации энергии и два круговорота — твердых и газообразных веществ. В структуре и функции экосистемы воплощены все виды активности организмов, входящих в данное биотическое сообщество: взаимодействия с физической средой и друг с другом. Однако организмы живут для самих себя, а не для того, чтобы играть какую-либо роль в эко- 140 системе. Свойства экосистемы слагаются благодаря деятельности входящих в нее растений и животных. Лишь учитывая это, мы можем понять ее структуру и функции, а также то, что экосистема как единое целое реагирует на изменения факторов среды. Проиллюстрируем данное положение на примере изменений, происходящих в сосновых лесах под действием сернистого ангидрида. 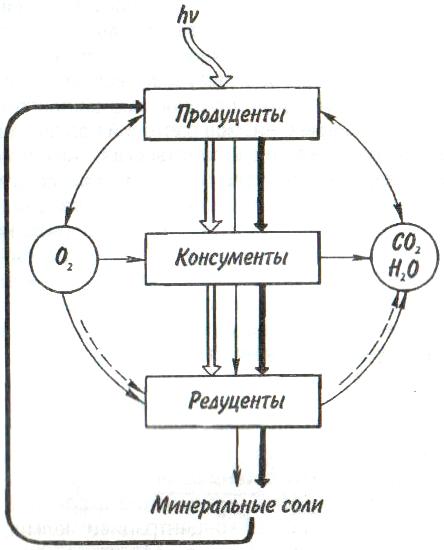 Рис. 8.4. Структура экосистемы, включающая один поток энергии (контурная стрелка) и два круговорота веществ: твердых (толстая стрелка) и газообразных (тонкая стрелка). Тонкой прерывистой стрелкой показано участие в круговороте анаэробных бактерий. Под действием сернистого газа в хвое сосен происходят значительные физиологические и морфометрические изменения. Наблюдается пожелтение концов хвоинок, а затем и их некроз, что в конечном итоге приводит к зна- 141 чительному уменьшению охвоенности, суховершинности и разреженности крон деревьев. Под влиянием кислых осадков отмечается обеднение травянисто-кустарникового яруса, появление множества мертвопокровных участков, что вызывает общее повышение температуры воздуха под пологом леса, в первую очередь в напочвенном ярусе. Длительная загазованность воздуха вызывает хроническое расстройство сосновых древостоев, замедляет их рост и ослабляет устойчивость не только к абиотическим факторам среды, но и к хвоегрызущим вредителям. Увеличению плотности хвоегрызущих чешуекрылых в зоне загрязнения способствуют ослабление физиолого-биохи-мических защитных механизмов растений под воздействием выбросов, содержащих сернистый газ, снижение биотического пресса на популяции вредителей со стороны паразитических насекомых, хищников и болезней. В целом сернистый газ отрицательно влияет на развитие хвоегрызущих чешуекрылых. Уменьшается масса гусеницы и куколки, ухудшаются репродуктивные показатели самок и жизнеспособность отложенных ими яиц. Однако плотность популяции этих насекомых увеличивается. Во-первых, снижается смертность куколок, так как в результате повышения температуры под пологом леса гусеницы успевают закончить развитие до того, как под воздействием заморозков осенью уйти в подстилку. Во-вторых, более чувствительные к загрязнению хищники и паразиты снижают свое давление на хвоегрызущих чешуекрылых. Кроме того, уменьшение охвоенности сопровождается еще большей концентрацией мелких, и без того многочисленных гусениц на хвое, что в итоге приводит к быстрой гибели сосновых лесов. |