Бродский А.К. Краткий курс общей экологии. Учебное пособие. Спб. Деан. 2000. 224 с
 Скачать 2.74 Mb. Скачать 2.74 Mb.
|

|
162 метаболизма имеют более высокие темпы размножения и обновления биомассы, т. е. более высокую продуктивность. 9.3. ТРОФИЧЕСКАЯ СТРУКТУРА ЭКОСИСТЕМЫ В результате рассеяния энергии в пищевых цепях и благодаря такому фактору, как зависимость метаболизма от размера особей, каждое сообщество приобретает определенную трофическую структуру, которую можно выразить либо числом особей на каждом трофическом уровне, либо урожаем на корню (на единицу площади), либо количеством энергии, фиксируемой на единице площади за единицу времени на каждом последующем трофическом уровне. Графически это можно представить в виде пирамиды, основанием которой служит первый трофический уровень, а последующие образуют этажи и вершину пирамиды. Различают три основных типа экологических пирамид — пирамиды чисел, биомассы и энергии. Пирамидачисел, или распределение особей по трофическим уровням, зависит от следующих факторов:
Для поддержания жизни одного льва требуется 50 зебр в год. На Фолклендских островах кулик-сорока на скалах во вре- 163 мя отлива добывает брюхоногого моллюска «блюдечко», но может справиться только с особями среднего размера; моллюски, размер которых превышает 45 мм, от него ускользают. Однако из этого правила есть исключения. Волки, охотясь сообща, могут убивать жертву более крупную, чем они сами, например оленей. Пауки и змеи, обладая свойством выделять яд, убивают крупных животных. Единственным же видом, способным использовать в пищу животных любой величины, является человек. В случае пастбищных пищевых цепей леса, когда продуцентами служат деревья, а первичными консументами — насекомые, уровень первичных консументов численно богаче особями уровня продуцентов. Таким образом, пирамиды чисел могут быть обращенными. Для примера на рис. 9.7 приведены пирамиды чисел экосистем степи и леса умеренной зоны. 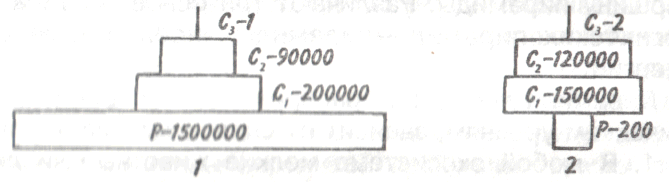 Рис. 9.7. Пирамида чисел для степи летом (1) и для леса умеренной зоны летом (2) (из Одума, 1975). Р —продуценты, С — консументы. Пирамидыбиомассыпредставляют более фундаментальный интерес, так как в них устранен «физический» фактор и четко показаны количественные соотношения биомасс. Если организмы не слишком сильно различаются по размеру, то, обозначив на трофических уровнях общую массу особей, можно получить ступенчатую пирамиду. Но если организмы низших уровней в среднем мельче организмов высших уровней, то имеет место обращенная пирамида биомассы. Например, в экосистемах с очень мелкими продуцентами и крупными консументами общая масса последних может быть в любой 164   данный момент выше общей массы продуцентов. Для пирамид биомассы можно сделать несколько обобщений. данный момент выше общей массы продуцентов. Для пирамид биомассы можно сделать несколько обобщений.
Пирамидаэнергиииз трех рассматриваемых типов экологических пирамид дает наиболее полное представление о функциональной организации сообщества, так как число и масса организмов, которые могут существовать на каждом трофическом уровне в тех или иных условиях, зависят не от количества фиксированной энергии, имеющейся в данное время на предыдущем уровне, а от скорости продуцирования пищи. В противоположность пирамидам чисел и биомассы, отражающим статику системы, т. е. характеризующим количество организмов или их биомассу в данный момент, пирамида энергии отра- 165 жает скорость прохождения массы пищи через пищевую цепь. На форму этой пирамиды не влияют изменения размеров особей и интенсивности их метаболизма, и если учтены все источники энергии, то пирамида всегда будет иметь «канонический» вид, как это диктуется вторым законом термодинамики. Для того чтобы оценить достоинства модели пирамиды энергии, сравним пирамиды биомассы и энергии одной из немногих экосистем, для которых известны все компоненты сообщества, включая редуцентов, — экосистемы ручьев Силвер-Спрингс во Флориде (рис.9.8). Продуценты в данной экосистеме представлены стрелолистом и прикрепленными водорослями. Среди кон- 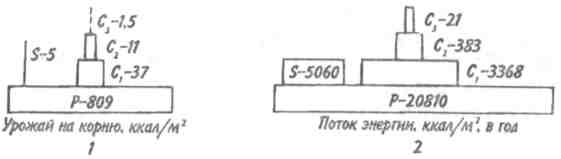 Рис. 9.8. Пирамиды биомассы (1) и энергии (2) в системе ручьев Силвер-Спрингс во Флориде (из Одума, 1975). S —редуценты. Р, С — см. рис. 9.7. сументов I порядка большое количество насекомых, брюхоногих моллюсков, растительноядных рыб и черепах. Различные виды рыб и хищные насекомые занимают следующий трофический уровень, а окунь, паразиты и панцирная щука - самый верхний уровень. Редуценты не только разлагают растительный материал, но уничтожают и органическое вещество других уровней: на пирамиде биомассы им соответствует линия, опирающаяся на первый трофический уровень. Биомасса бактерий и грибов очень мала по сравнению с их ролью в энергетическом потоке сообщества. Поэтому в пирамиде чисел значение редуцентов сильно преувеличено, а в пирамиде биомассы сильно преуменьшено. Исходя из данного положения, сформулируем «экологическое правило»: данные по численности приводят к преувеличению зна- 166    чения мелких организмов, а данные по биомассе — к преувеличению роли крупных организмов. чения мелких организмов, а данные по биомассе — к преувеличению роли крупных организмов.Следовательно, эти критерии непригодны для сравнения функциональной роли популяций, сильно различающихся по значению отношения интенсивности метаболизма к размеру особей, хотя, как правило, биомасса более надежный критерий, чем численность. В то же время поток энергии служит более подходящим показанием при сравнении компонента с любым другим и экосистемы в целом с соседней экосистемой. Поток энергии в экосистеме представляет интерес не только для эколога, но и для каждого из нас. Объясняется это тем, что, уяснив основные законы превращения энергии, мы лучше поймем, почему природные экосистемы функционируют именно так, как они функционируют. Не менее важно и то, что такое знание поможет нам определить границы, за которыми наше воздействие на окружающую среду принесет ей непоправимый ущерб. 167 Те м а 10 БИОГЕОХИМИЧЕСКИЕ ЦИКЛЫ Распределение энергии — не единственное явление, обусловленное пищевыми цепями. Некоторые вещества по мере продвижения по цепи не рассеиваются, а, наоборот, накапливаются. 10.1. ОБМЕННЫЙ И РЕЗЕРВНЫЙ ФОНДЫ Известно, что из более 90 химических элементов, встречающихся в природе, 30-40 необходимы живым организмам. Некоторые элементы, такие как углерод, водород и азот, требуются в больших количествах, другие в малых или даже минимальных количествах. Какова бы ни была потребность в них, все элементы участвуют в биогеохимических круговоротах. Биогеохимический круговоротимеет вид кольца, направленного от авто- 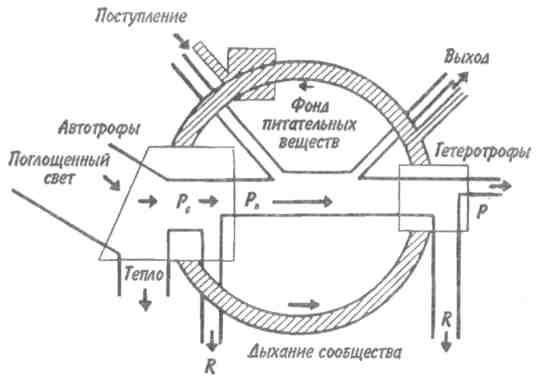 Рис. 10.1. Биогеохимический круговорот (заштрихованное кольцо) на фоне упрощенной схемы потока энергии (по Одуму, 1975). Обозначения см. на рис. 9.4. 168      трофов к гетеротрофам и от них снова к автотрофам (рис. 10.1). трофов к гетеротрофам и от них снова к автотрофам (рис. 10.1).В природе элементы никогда или почти никогда не бывают распределены равномерно по всей экосистеме и находятся всюду в разной химической форме. На пути между гетеротрофами и автотрофами элементы попадают в так называемый резервный фонд. Резервныйфонд— большая масса медленно движущихся веществ, в основном не связанных с организмами. В отличие от него, обменныйфондпредставляет собой быстрый обмен между организмами и их непосредственным окружением и имеет вид кольца. В зависимости от природы резервного фонда выделяют два основных типа биогеохимических круговоротов: 1) круговорот газообразных веществ с резервным фондом в атмосфере или гидросфере, 2) осадочный цикл с резервным фондом в земной коре. Резервные фонды в атмосфере и гидросфере легко доступны, поэтому такие биогеохимические круговороты относительно устойчивы. Осадочные циклы, в которых участвуют фосфор и железо, гораздо менее стабильны. Они более подвержены влиянию различного рода местных изменений, так как основная масса вещества сосредоточена в малоактивном и малоподвижном резервном фонде земной коры. Следовательно, если «спуск», т.е. поступление веществ из обменного фонда в резервный, совершается быстрее, чем «подъем», то часть обмениваемого материала выходит из круговорота. Обменный фонд образуется за счет веществ, которые возвращаются в круговорот двумя основными путями: 1) в результате первичной экскреции животными и 2) при разложении детрита микроорганизмами. Если оба пути замыкания обменного фонда реализуются в одной экосистеме, то первый из них доминирует, например, в планктоне и других сообществах, где основной поток энергии идет через пастбищную пищевую цепь; второй путь преобладает в степях, лесах умеренной зоны и других сообществах, в которых основной поток энергии направлен через детритную пищевую цепь. 169 10.2. БЛОЧНАЯ МОДЕЛЬ КРУГОВОРОТА Существуют различные способы изображения биогеохимических круговоротов. Выбор способа зависит от особенностей биогеохимического цикла того или иного элемента. При обсуждении круговорота кислорода экологи обычно различают пути, связанные с химическим включением кислорода в органические соединения, и пути, сопряженные с передвижением воды. Круговорот воды, или гидрологический цикл, хорошо сбалансирован в масштабе земного шара и приводится в движение энергией, в основном не связанной с организмами. Особи быстро теряют воду путем испарения и выделения; за время жизни особи содержащаяся в организме вода может обновляться сотни и тысячи раз. В то же время участие организмов в обмене воды ничтожно мало — общий объем испарения и транспирации оценивается в 59⋅1018 г в год, в связи с чем при изображении биогеохимического цикла воды 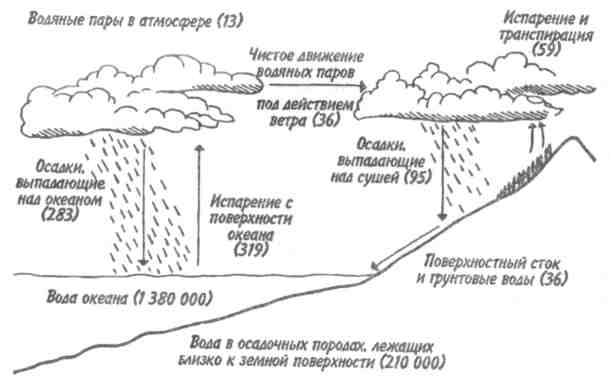 Рис. 10.2. Круговорот воды и его главные компоненты в глобальном масштабе (по: Риклефс, 1979). Цифры в скобках-количество воды, миллиарды миллиардов (1018) граммов в год. 170  делают акцент на резервном, а не на обменном фонде (рис. 10.2). делают акцент на резервном, а не на обменном фонде (рис. 10.2).При изображении биогеохимических циклов других веществ делают акцент на обмене между организмами и резервным фондом, а также на путях движения веществ внутри экосистемы. Так, круговорот углерода и кислорода обеспечивается комплементарными процессами фотосинтеза и дыхания. Азот, фосфор и сера проделывают в экосистеме более сложный путь, причем в этом им помогают микроорганизмы со специализированными метаболическими функциями. Любую экосистему можно представить в виде ряда блоков, через которые проходят различные материалы и в которых эти материалы могут оставаться на протяжении различных периодов времени (рис. 10.3). В круговоротах минеральных веществ в экосистеме, как правило, участвуют три активных блока: живые организмы, мертвый органический детрит и доступные неорганические вещества. Два добавочных блока — косвенно доступные неорганические вещества и осаждающиеся органические вещества — связаны с круговоротами биогенных элементов в каких-то периферических участках общего цикла (рис. 10.3), однако обмен между этими блоками и остальной экосистемой замедлен по сравнению с обменом, происходящим между активными блоками. Процессы ассимиляции и распада, благодаря которым происходят круговороты биогенных элементов в биосфере, тесно связаны с поглощением и освобождением энергии организмами. Следовательно, пути биогенных элементов параллельны потоку энергии через сообщество. В наибольшей степени с превращениями энергии в сообществе связан круговорот углерода, так как большая часть энергии, ассимилированной в процессе фотосинтеза, содержится в органических углеродсодержащих соединениях. В результате процессов, сопровождающихся выделением энергии, среди которых самым главным является дыхание, углерод высвобождается в 171 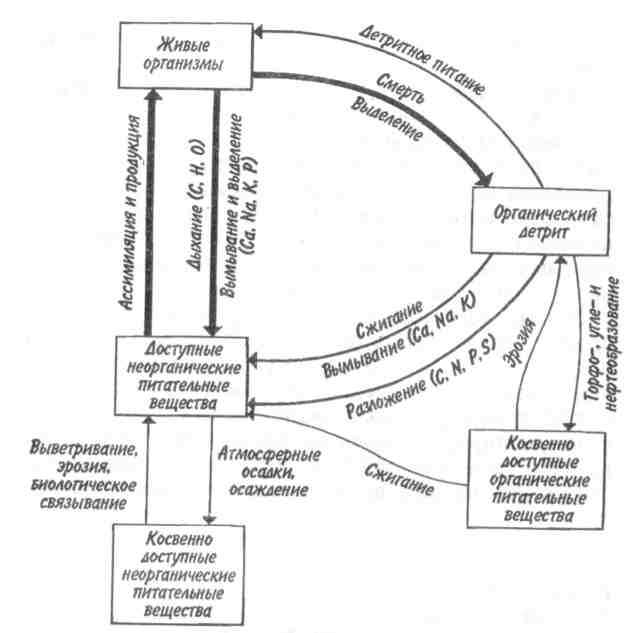 Рис. 10.3. Блочная модель экосистемы с указанием некоторых наиболее важных путей обмена минеральных веществ (по: Риклефс, 1979). виде углекислого газа. Когда в организме происходит метаболизм органических соединений, содержащих азот, фосфор и серу, последние нередко удерживаются в этом организме, поскольку они необходимы для синтеза структурных белков, ферментов и других органических молекул, образующих структурные и функциональные компоненты живых тканей. А поэтому прохождение азота, фосфора и серы через каждый трофический уровень несколько замедленно по сравнению со средним временем переноса энергии. 172     10.3. ПРИМЕРЫ НЕКОТОРЫХ БИОГЕОХИМИЧЕСКИХ ЦИКЛОВ 10.3. ПРИМЕРЫ НЕКОТОРЫХ БИОГЕОХИМИЧЕСКИХ ЦИКЛОВКаждый химический элемент, совершая круговорот в экосистеме, следует по своему особому пути, но все круговороты приводятся в движение энергией, и участвующие в них элементы попеременно переходят из органической формы в неорганическую и обратно. Рассмотрим круговороты некоторых химических элементов с учетом особенностей поступления их из обменного фонда в резервный и возврата в обменный фонд. Биогеохимический цикл азота — пример очень сложного круговорота вещества с резервным фондом в атмосфере (рис. 10.4). Азот, входящий в состав белков и других азотсодержащих соединений, переводится из 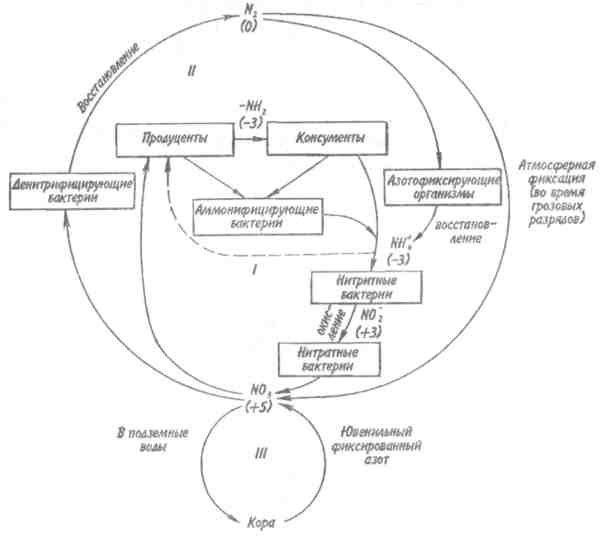 Рис. 10.4. Биогеохимический цикл азота. Здесь и на рис. 10.5-10.7: / — обменный фонд; //, ///—резервные фонды. 173 органической формы в неорганическую в результате деятельности ряда бактерий — редуцентов, причем каждый вид бактерий выполняет свою часть работы. Особенность биогеохимического цикла фосфора (рис. 10.5) состоит в том, что редуценты переводят фосфор из органической формы в неорганическую, не окисляя его. Цикл фосфора менее совершенен, чем цикл азота, так как в результате происходит утечка этого элемента в глубокие осадки. Биогеохимический цикл серы характерен обширным резервным фондом в земной коре, и меньшим — в ат- 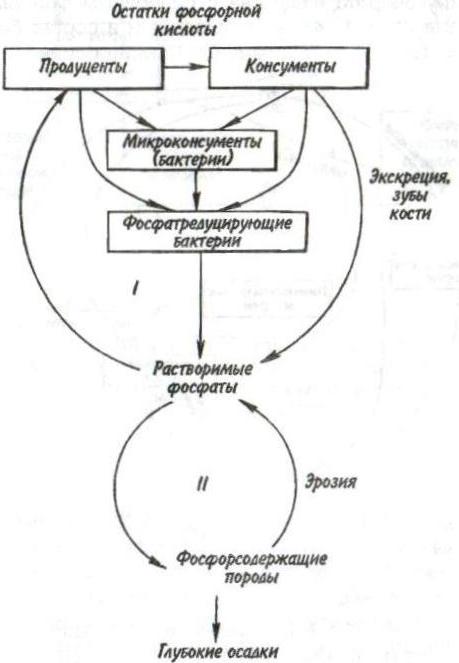 Рис. 10.5. Биогеохимический цикл фосфора. 174 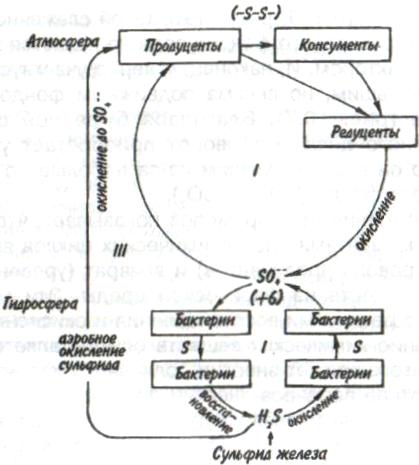 Рис. 10.6. Биогеохимический цикл серы. 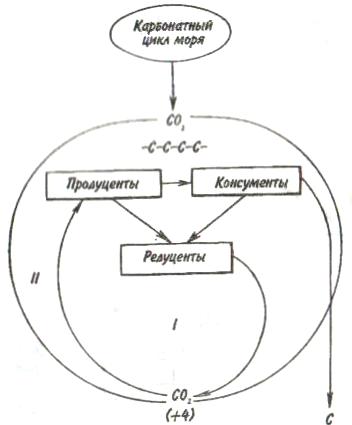 Рис. 10.7. Биогеохимический цикл углерода. 175 мосфере (рис. 10.6). В результате такой слаженности обменного и резервного фондов сера не является лимитирующим фактором. И, наконец, углерод участвует в цикле с небольшим, но весьма подвижным фондом в атмосфере (рис. 10.7). Благодаря буферной системе карбонатного цикла круговорот приобретает устойчивость, но он все-таки уязвим из-за небольшого объема резервного фонда (0,029% С02). Рассмотрение этих примеров показывает, что критическими моментами биогеохимических циклов являются захват (уровень продуцентов) и возврат (уровень редуцентов) веществ из физической среды. Эти моменты связаны с реакциями восстановления и окисления. Восстановление химических веществ осуществляется в конечном итоге за счет энергии солнечного излучения. На каждом этапе переноса энергии происходит ее рассеивание, заканчивающееся на уровне редуцентов, которые окисляют элементы до состояния, в котором они уже могут быть захвачены продуцентами. В целом на уровне обменного фонда биогеохимический круговорот может быть представлен системой ступенек, в пределах каждой из которых осуществляется своя часть процесса окисления (рис. 10.8). Таким образом, важнейшее свойство потоков в экосистемах—их цикличность. Вещества в экосистемах со- 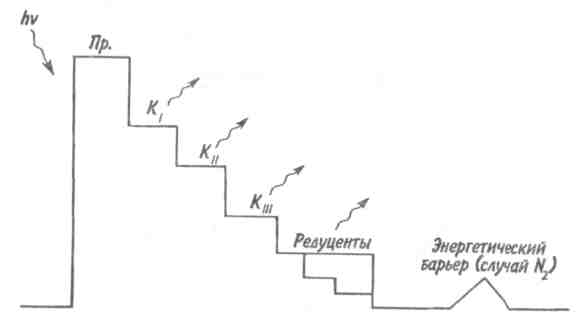 Рис. 10.8. Принцип движения веществ в обменном фонде. 176 вершают практически полный круговорот, попадая сначала в организмы, затем в абиотическую среду и вновь возвращаясь к организмам. В круговоротах участвуют не только биогенные элементы, но и многие загрязняющие вещества. Некоторые из них не только циркулируют в окружающей среде, но и имеют тенденцию накапливаться в организмах. В таких случаях концентрация какого-либо загрязняющего вещества, обнаруженного в организмах, нарастает по мере прохождения его вверх по пищевой цепи, гак как организмы быстрее поглощают загрязняющие вещества, чем выделяют их. Ртуть, например, может содержаться в воде и придонном иле в относительно безвредных концентрациях, тогда как ее содержание в организме водных животных, имеющих раковину или панцирь, может достигать летального для них уровня. Действие пестицидов, таких как ДДТ, основывается на сходном принципе: содержание их в воде может быть столь незначительным, что выявить их практически не удается, однако чем выше трофический уровень, на котором находится данный организм, тем больше концентрация пестицида в его тканях. Это явление известно под названием биологического усиления, или биологического накопления. 177 Тема 11 БИОТИЧЕСКОЕ СООБЩЕСТВО На примере биогеохимических циклов мы рассмотрели (см. рис. 9.2) связь между биотическими и абиотическими компонентами экосистемы. Биотическая часть называется также биотой, биоценозом или биотическим сообществом. Биотическое сообщество — это любая совокупность популяций, населяющих определенную территорию (биотоп), это своего рода организационная единица в том смысле, что она обладает некоторыми особыми свойствами, не присущими слагающим ее компонентам — особям и популяциям. Биотическое сообщество постоянно меняет внешний облик (мысленно сравните осенний и зимний лес), при этом оно обладает собственной структурой и функциями. |