Бродский А.К. Краткий курс общей экологии. Учебное пособие. Спб. Деан. 2000. 224 с
 Скачать 2.74 Mb. Скачать 2.74 Mb.
|

|
12.1. ЭКОЛОГИЧЕСКАЯ СУКЦЕССИЯ Автогенные изменения называют развитием экосистемы, или экологической сукцессией. При определении экологической сукцессии следует учитывать три момента:
Скорость изменений и время, необходимое для достижения стабилизированного состояния, варьируют в разных экосистемах и для разных признаков экосистемы. Кривая изменения общей биомассы обычно имеет выпуклую форму, тогда как кривая изменения валовой первичной продукции — вогнутую (рис. 12.1). Как в лесной экосистеме, так и в условиях лабораторного микрокосма чистая первичная продукция возрастает на ранних стадиях развития сообщества, но убывает — на поздних. Оценка изменений этих и других параметров приведена в табл. 12.1. 189 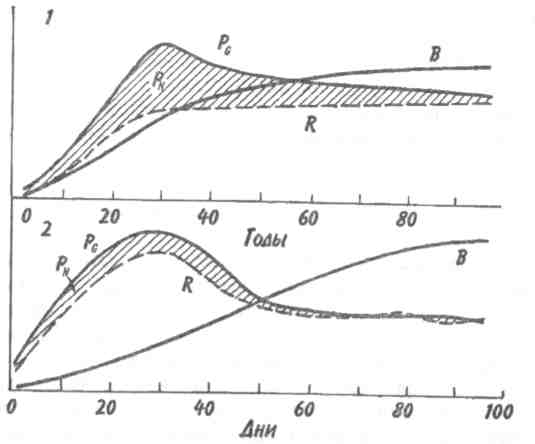 Рис. 12.1. Энергетика развития экосистемы леса (/) и лабораторного микрокосма (2) (по: Одум, 1975). Остальные обозначения см. на рис. 9.4. В — общая биомасса. На ранних стадиях экологической сукцессии валовая первичная продуктивность превосходит затраты на дыхание сообщества, так что Р/R>1. Для особого случая, т. е. для случая сильного органического загрязнения среды (например, в сточных водах), затраты на дыхание могут превалировать над валовой первичной продуктивностью. Если сукцессия начинается с подобного состояния, то говорят о гетеротрофнойсукцессии в отличие от обычной, авто-трофнойсукцессии. Однако в любом случае по мере развития экосистемы отношение P/Rстремится к 1. Графически это можно изобразить в виде прямой, составляющей угол 45 ° с осями координат (рис. 12.2), несмотря на то, что в разных экосистемах, в зависимости от физических факторов среды, абсолютные значения продуктивности и, следовательно, затрат на дыхание сообщества могут существенно различаться. Пока Рбольше R, в системе будут накапливаться органическое вещество и биомасса, в результате чего отно- 190  Рис. 12.2. Положение сообществ различных типов в классификации, основанной на метаболизме (по: Одум, 1975). Направления автотрофной и гетеротрофной сукцессии указаны стрелками. Сообщества, расположенные вдоль диагонали, потребляют за год в среднем примерно столько, сколько они создали, и их можно считать метаболически климаксными. шение Р/Вбудет снижаться, а отношения В/Р, B/Rили В/Е (где Е= Р+ R) соответственно увеличиваться. Иными словами, биомасса, поддерживаемая доступным потоком энергии, достигает максимума в зрелых экосистемах (табл. 12.1, признак 3). Как следствие этого, чистая продукция сообщества — урожай за годовой цикл — велика на ранних стадиях развития экосистемы и мала или равна нулю в зрелом ее состоянии (признак 4). По мере развития экосистемы следует ожидать тонких изменений в структуре пищевых цепей (признак 5). При ненарушаемом течении сукцессии имеется достаточно времени для развития более тесных связей и взаимных адаптации между животными и растениями, что ведет к 191 появлению множества механизмов, уменьшающих выедание растений: образуются плохо перевариваемые опорные ткани, увеличивается давление хищников на фитофагов и т. д. Листва опадает, и переработка органического вещества переходит из зеленого пояса в коричневый.  192  Важной тенденцией в развитии экосистемы является стремление к замыканию биогеохимических круговоротов основных элементов, таких как азот, фосфор и кальций. Зрелые системы по сравнению с развивающимися обладают большей способностью захватывать вещества и сохранять их в обменном фонде. Они теряют лишь очень небольшие количества веществ по сравнению с незрелыми или нарушенными системами. Отсюда следуют изменения, отмеченные в табл. 12.1 (признаки 6-8). Важной тенденцией в развитии экосистемы является стремление к замыканию биогеохимических круговоротов основных элементов, таких как азот, фосфор и кальций. Зрелые системы по сравнению с развивающимися обладают большей способностью захватывать вещества и сохранять их в обменном фонде. Они теряют лишь очень небольшие количества веществ по сравнению с незрелыми или нарушенными системами. Отсюда следуют изменения, отмеченные в табл. 12.1 (признаки 6-8).В развивающихся системах мало число видов и мала их выравненность, не развиты гетеротипические реакции, а в зрелых - число видов и их выравненность велики, гетеротипические реакции развиты (признаки 9 -11). Особенно примечательны в этом отношении тесные связи между неродственными видами, например между кораллами (кишечнополостные) и зооксантеллами (одноклеточные жгутиковые) или между микоризой и деревьями. Как следствие усиления гетеротипических реакций происходит специализация по нишам и усиление стратификации (признаки 12, 13). Рассмотрим подробнее признаки 14 и 15. На начальной стадии сукцессии организмы обычно бывают мелкими, для них характерны простые жизненные циклы и высокие скорости размножения. Изменения размеров особей происходят вследствие перемещения биогенных веществ из неорганической в органическую фазу. Малые размеры дают преимущество, особенно автотрофам, в среде, богатой минеральными веществами, поскольку возрастает отношение площади поверхности тела к его объему. Однако, по мере развития экосистемы, неорганические биогенные вещества становятся все более связанными в биомассе, так что преимущество переходит к организмам с большим объемом, т. е. к более крупным — либо к более крупным особям данного вида, либо к видам с более крупными особями, либо к тем и другим, так или иначе обладающим большими возможностями для накопления веществ и более сложными жизненными циклами. 193 Характер роста популяции меняется от экспоненциального к логистическому, так как возрастает сопротивление среды. Известно, что в ограничении роста популяции, когда увеличение ее численности происходит по логистическому закону, решающее значение имеет отрицательная обратная связь. Во многих случаях биотическая регуляция выедания растительности (признак 5), плотности популяции (признак 16) и круговорота веществ (признак 6) служит основным механизмом действия отрицательной обратной связи. В результате расширения поля деятельности для отрицательной обратной связи увеличивается стабильность экосистемы (признак 17), снижается ее энтропия (признак 18) и в конечном итоге повышается количество содержащейся в ней информации (признак 19). При загрязнении или иной форме стресса вектор изменения признаков экосистемы становится противоположным. В этом легко убедиться, анализируя изменения, которые сопровождают значительное загрязнение нефтепродуктами малых рек Западной Сибири. Это обеднение видового состава во всех звеньях пищевых цепей (22 вида в фитопланктоне вместо 31; 4 вида, высших растений вместо 28; 1 вид ветвистоусых ракообразных в зоопланктоне вместо 5; 20 видов в бентосе вместо 45 и т. д.), концентрация доминирования в немногих видах, устойчивых к загрязнению, уменьшение стратификации (слоистости) сообщества, т. е. равномерности распределения организмов по микробиотопам, уменьшение суммарной биомассы (либо в результате снижения видового разнообразия, либо в результате того, что особи видов, устойчивых к нефтяному загрязнению, имеют меньший индивидуальный вес), сужение кормовой базы рыб вследствие угнетения зоопланктона, животных-—обитателей зарослей высших водных растений, и концентрации биомассы в мезобентосе. Таким образом, знание изменений, которые происходят в процессе экологической сукцессии, во многих случаях открывает возможность для прогноза неблагоприятных последствий антропогенного воздействия на экосистемы. Основной результат деятельности сообщества состоит в усилении гетеротипических реакций, сохранении 194 минеральных веществ, повышении стабильности и увеличении информации. Этот результат называется всеобщим гомеостазом. Стратегия экологической сукцессии направлена на достижение такой обширной и разнообразной органической структуры, какая только возможна в границах, установленных доступным потоком энергии и физическими условиями существования. 12.2. ПЕРВИЧНАЯ И ВТОРИЧНАЯ СУКЦЕССИИ. ПОНЯТИЕ КЛИМАКСА Экологическая сукцессия протекает через ряд этапов, при этом биотические сообщества сменяют друг друга. Замещение видов в сукцессии вызвано тем, что популяции, стремясь модифицировать окружающую среду, создают условия, благоприятные для других популяций. Это продолжается до тех пор, пока не будет достигнуто равновесие между биотическими и абиотическими компонентами. Последовательность сообществ, сменяющих друг друга в конкретном районе, называется серией, причем лишь немногие виды сохраняются от начальных этапов сукцессии до зрелого состояния экосистемы. Если развитие экосистемы начинается на участке, который перед этим не был занят каким-либо сообществом (недавно вышедшая на поверхность скала, песок или лавовый поток), процесс называется первичной сукцессией. Если развитие экосистемы происходит на площади, с которой удалено предыдущее сообщество (например, заброшенное поле или вырубка), то это будет вторичная сукцессия. Она протекает обычно быстрее первичной, поскольку на территории, которая ранее была занята, уже имеются некоторые организмы, необходимые для обмена веществ со средой, более благоприятной для развития сообщества, чем «стерильная» зона. Примером первичной сукцессии может служить зарастание песчаных дюн озера Мичиган. Сообщество первых поселенцев (пионеры) на дюнах состоит из злаков, ивняка, виш- 195 ни, хлопкового дерева и животных, таких как жуки-скакуны, норные пауки и кузнечики. За сообществом первых поселенцев следуют лесные сообщества, каждое из которых обладает своим животным миром. Несмотря на то, что развитие началось на очень сухом и бесплодном месте, в конечном итоге здесь появился буково-кленовый лес, в отличие от голых дюн влажный и холодный. Мощная, богатая гумусом почва с дождевыми червями и моллюсками в лесу контрастирует с сухим песком, на котором она образовалась. В качестве примера вторичной сукцессии приведем восстановление елового леса. После вырубки или пожара условия на месте ельника настолько изменяются, что ель не может снова заселить освободившуюся площадь. На открытых местах всходы ели повреждаются поздними весенними заморозками, страдают от перегрева и не могут конкурировать со светолюбивыми растениями. В первые два года на вырубках и гарях буйно развиваются травянистые растения: кипрей, вейник и др. Вскоре появляются многочисленные всходы березы, осины, а иногда сосны, семена которых легко разносятся ветром. Деревья вытесняют травянистую растительность и постепенно образуют мелколистный или сосновый лес. Только тогда возникают условия, благоприятные для возобновления ели. Теневыносливые всходы ели успешно конкурируют с подростом светолюбивых лиственных пород. Когда ель достигает верхнего яруса, она полностью вытесняет лиственные деревья. В принципе так же протекает сукцессия пихтово-кедровой тайги (рис. 12.3). Каждый последующий этап сукцессии длится дольше предыдущего, характеризуется более высоким значением отношения биомассы к единице потока энергии (В/Р + R) и своими видами-доминантами. Особенно сильное воздействие на среду оказывают доминирующие виды растений. Большой вклад растений в формирование сообщества связан не только с их ролью первичных продуцентов, но и с тем, что они медленно разлагаются. Растения образуют не только биомассу, но и основную часть некромассы, т. е. мертвого органического вещества. Несмотря на высокую активность бактерий и детритофагов, растительные остатки накаплива- 196 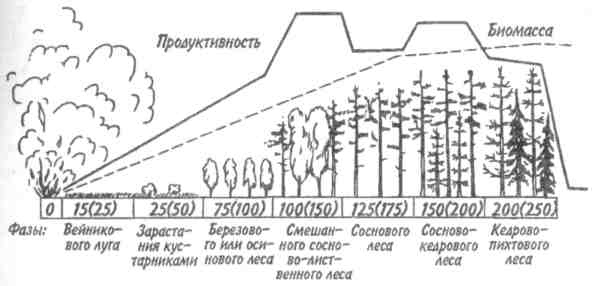 Рис. 12.3. Сукцессия сибирского темнохвойного леса (пихто- во-кедровая тайга) после опустошительного пожара (по Реймерсу, 1990). Числа показывают колебания длительности (в годах) прохождения фаз сукцессии (в скобках указан срок их окончания). Биомасса и биологическая продуктивность даны в произвольном масштабе. ются в виде листового опада или торфа. Кроме того, доминирование деревьев в огромном числе сообществ объясняется именно их способностью накапливать в составе организма мертвое вещество, из которого состоит большая часть ствола и ветвей. Способность кустарников и деревьев в умеренно влажных местообитаниях вытеснять травяную растительность в значительной степени связана с развитием у них кроны (и корневой системы) на протяженном и сложноразветвленном «скелете», состоящем в основном из мертвой опорной ткани (древесины). В свою очередь сукцессия в наземных местообитаниях влечет за собой регулярную смену растительных форм. Растения, обнаруживаемые на ранних и поздних стадиях сукцессии, характеризуются различными стратегиями роста и размножения (табл. 12.2). Растения, относящиеся к ранним стадиям сукцессии, благодаря своей высокой способности к расселению быстро занимают вновь образовавшиеся или нарушенные местообитания. Позднесукцессионные виды распространяются и растут медленнее, но теневыносливость их подростков и большие размеры зрелых растений дают им преимущества в конкуренции с видами, образующими ранние стадии сукцессии. Растения терминальных сообществ приспособлены к росту и процветанию в той среде, которую созда- 197 ют они сами, тогда как виды, появляющиеся на ранних стадиях сукцессии, обладают способностью к колонизации еще не используемых сред. Тела животных разлагаются гораздо быстрее, однако иногда их остатки, как и остатки растений, определяют структуру сообщества и ход сукцессии в нем. Это имеет место, например, при накоплении обызвествленных скелетов в процессе роста кораллов. Чаще животные пассивно реагируют на сукцессию растительности. Не исключено, конечно, что семено-ядные птицы также влияют на смену растительности. Таблица 12.2. Общие характеристики растений, свойственных разным стадиям сукцессии (по Риклефсу, 1979) 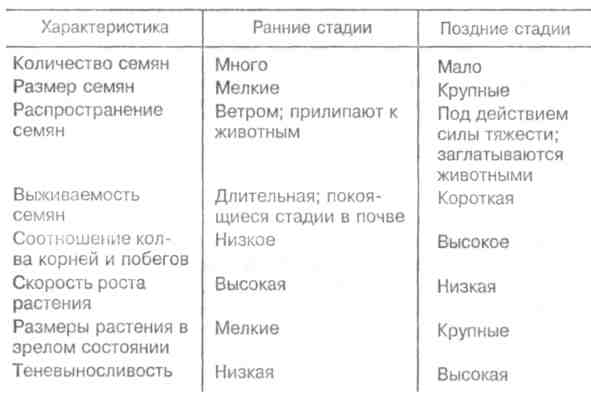 Терминальным, или стабильным, сообществом развивающейся серии является климаксноесообщество. В климаксном сообществе, в отличие от сообществ развивающихся и других нестабильных стадий, минимальна или полностью отсутствует годовая чистая продукция органического вещества. Для каждой климатической зоны удобно различать единственный климатический климакс и различное число здафических климаксов. Климатическийклимакс — это теоретическое сообщество, к достижению которого направлено все развитие экосис- 198  темы в данном районе, находящееся в равновесии с общими климатическими условиями. Теоретическое сообщество реализуется там, где физические условия среды не столь экстремальны, чтобы изменять воздействие преобладающего климата. Там, где рельеф местности, почва, водоемы, пожары, заболачивание и другие факторы препятствуют развитию климатического климакса, сукцессия заканчивается формированием эдафического климакса. Так, в зависимости от рельефа и особенностей почвы на примыкающих друг к другу морских террасах с одинаковой материнской породой развиваются различные сообщества (рис. 12.4). Поскольку основным модифицирующим фактором экосистемы является биотическое сообщество, то чем более экстремальны физические условия среды, тем больше вероятность того, что развитие экосистемы прекратится, не достигнув равновесия с общими климатическими условиями. темы в данном районе, находящееся в равновесии с общими климатическими условиями. Теоретическое сообщество реализуется там, где физические условия среды не столь экстремальны, чтобы изменять воздействие преобладающего климата. Там, где рельеф местности, почва, водоемы, пожары, заболачивание и другие факторы препятствуют развитию климатического климакса, сукцессия заканчивается формированием эдафического климакса. Так, в зависимости от рельефа и особенностей почвы на примыкающих друг к другу морских террасах с одинаковой материнской породой развиваются различные сообщества (рис. 12.4). Поскольку основным модифицирующим фактором экосистемы является биотическое сообщество, то чем более экстремальны физические условия среды, тем больше вероятность того, что развитие экосистемы прекратится, не достигнув равновесия с общими климатическими условиями.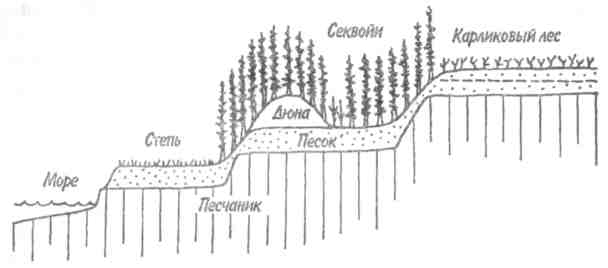 Рис. 12.4. Эдафические климаксы на прибрежном участке в Северной Калифорнии (по: Одум, 1975). Существует еще такое понятие, как катастрофический (или циклический) климакс. Катастрофическийклимакс—терминальное состояние экосистемы, возникающее благодаря периодически появляющимся катастрофам, таким как пожары, вредители и т. д. Природные катастрофы уничтожают климаксное сообщество и способствуют тем самым повторению сукцессии, длящейся до климаксного состояния, при котором экосистема особенно подвержена воздействию катастрофическо- 199 го фактора. Подобный процесс повторяется неоднократно, и со временем возникает естественная самоподдерживающаяся система, для существования которой необходимы регулярно возникающие катастрофы. Иногда сообщества катастрофического климакса занимают большие территории. Примером могут служить заросли чапарраля в Калифорнии, покрывающие площади до 2—3 млн га. В данном случае результатом сукцессии является кустарниковое сообщество, особенно подверженное пожарам. Существует представление, что любая сукцессия — это циклический процесс. Цикличность изменений может быть весьма различной и подчас очень сложной в связи с наложением циклов разной длительности друг на друга. В некоторых случаях те или иные фазы цикла сохраняются неопределенно долго, в связи с чем процесс не выглядит циклическим. Однако это не всегда так. Возьмем классический пример. Гарь зарастает мхами и пионерной травянистой растительностью, затем их сменяет ассоциация с малиной и кипреем, далее березняк, а затем неопределенно долго хвойный лес. Тем не менее, цикличность здесь очевидна: хвойный лес (особенно в более северных районах его произрастания) постепенно истощает почву, в связи с этим усиливается покров сфагновых мхов (не нуждающихся в почвенном минеральном питании) и, в конце концов, на этом месте остается (опять же неопределенно долгий срок) верховое болото. При чрезмерном росте болота сфагновый покров редеет, открытая поверхность торфа начинает пылить, торф самовозгорается и после пожара остается гарь, обогащенная минеральными элементами, накопленными как в мертвых остатках леса, так и в торфе. В итоге все может начинаться сначала. Человек часто влияет на развитие экосистемы, препятствуя достижению ею климаксного состояния. Когда сообщество, не представляющее собой климатический или эдафический климакс для данной местности, поддерживается человеком или домашними животными, то его называют дисклимаксомили антропогеннымсубкли- 200 максом. Например, чрезмерный выпас может породить пустынное сообщество там, где по условиям регионального климата могла бы сохраняться степь. Пустынные сообщества в данном случае—дисклимакс, а степь — климатический климакс. Итак, сукцессия — направленное развитие экосистемы, результатом которого является установление равновесия между биотическим сообществом и физической средой. В процессе сукцессии видовые популяции организмов и типы функциональных связей между ними закономерно, периодически и обратимо сменяют друг друга. В отличие от сукцессии эволюция экосистем представляет собой длительный процесс их исторического развития. Эволюционные процессы принципиально отличаются от описанных выше тем, что они необратимы, а следовательно, и ацикличны. При этом обычно необратимо одновременно меняются как макро-, так и микрохарактеристики системы. По отношению к биосфере в целом это, по сути дела, история развития жизни на Земле — от ее возникновения до наших дней. |