В. Н. Сайтаниди Рецензент членкорреспондент расхн в. Ф. Красота Петухов В. Л. и др. П31 Ветеринарная генетика В. Л. Петухов, А. И. Жигачев, Г. А. Назарова. 2е изд., перераб и доп. М. Колос, 1996. 384 с ил. Учебники
 Скачать 5.3 Mb. Скачать 5.3 Mb.
|

|
Глава 4 ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ СЦЕПЛЕННОЕ НАСЛЕДОВАНИЕ ПРИЗНАКОВ Мы познакомились с дигибридным скрещиванием и уяснили, Ч независимое комбинирование признаков объясняется тем, что расщепление одной пары аллельных генов, определяющих соответствующие признаки, происходит независимо от другой пары. Однако это наблюдается только в том случае, когда гены разных пар находятся в разных парах хромосом и при образовании половых клеток гибрида в мейозе отцовские и материнские хромосомы независимо комбинируются. Но количество хромосом очень ограниченно по сравнению с количеством признаков, каждый из которых развивается под контролем определенного гена. Так, у дрозофилы известно около 7000 генов при четырех парах хромосом. Предполагается, что у человека не менее 5Q тыс. генов при 23 парах хромосом, и т. д. Отсюда следует, что 3 каждой паре хромосом должны быть локализованы сотни аллелей. Естественно, что между генами, которые находятся в одной хромосоме, наблюдается сцепление и при образовании половых клеток они должны передаваться вместе. Сцепленное наследование открыли в 1906 г. английские генетики У. Бэтсон и Р. Пеннет при изучении наследования признаков у душистого горошка, но они не смогли вскрыть причины этого явления. Природу сцепленного наследования в 1910 г. выяснили ученые Т. Морган и его сотрудники К. Бриджес и А. Стертевант. В качестве объекта исследования они избрали плодовую муху дрозофилу, которая оказалась очень удобной для генетических опытов. В клетках тела дрозофилы находится 4 пары хромосом. Она отличается очень высокой плодовитостью — одна пара дает более ста потомков. У нее большая скорость развития — в течение 12—15 дней после оплодотворения из яйца развиваются личинка, куколка и взрослая особь, которая почти сразу же способна давать потомство. Можно исследовать в течение года более двадцати поколений. Мухи серого цвета, с красными глазами, имеют маленькие размеры (около 3 мм), легко разводятся в биологических пробирках; для изучения их признаков можно пользоваться лупами. При просмотре сотен тысяч особей Морган обнаружил множество разных мутаций: встреча- 51 лись мухи с черным и желтым телом, с белыми и другого цвета глазами, с измененной формой и положением крыльев и т. д. Иногда попадались особи, имеющие сразу несколько мутаций, например черное тело, зачаточные крылья, киноварные глаза. Изучая наследование разных пар признаков при дигибридном и полигибридном скрещиваниях, Морган и его сотрудники обнаружили большое число примеров сцепленного (совместного) их наследования. Все изученные признаки распределились на четыре группы сцепления в соответствии с числом и размерами хромосом у дрозофилы. На этом основании Морган сделал вывод о том, что гены, определяющие эти признаки, находятся в хромосомах. Гены, расположенные в одной хромосоме, представляют собой группу сцепления. Сцепление генов — это совместное наследование генов, расположенных в одной и той же хромосоме. Количество групп сцепления соответствует гаплоидному числу хромосом. Например, у дрозофилы 4 группы сцепления, у человека 23, у крупного рогатого скота 30, у свиней 19 и т. д. Мысль о расположении генов в хромосомах высказал Сеттон еще в 1902 г. Он обнаружил параллелизм в поведении хромосом в мейозе и наследовании признаков у одного из видов кузнечика. Дальнейшие исследования, проведенные Морганом, показали, что сцепление генов, расположенных в одной хромосоме, может быть полным или неполным. Наиболее четко разница в поведении сцепленных и независимо наследующихся генов выявляется при проведении анализирующего скрещивания. При независимом наследовании двух пар признаков у гибрида Fi (AaBb) с равной вероятностью образуется 4 сорта гамет: АВ, АЪ, аВ, аЪ. При скрещивании с полным рецессивом (aabb) количество сортов гамет у гибрида обусловливает число типов потомков и одинаковую вероятность их появления, так как гаметы рецессивной особи (ab) не могут изменить проявления доминантных и рецессивных генов гамет гибрида. В результате соотношение фенотипов потомства будет равно 1:1:1:1. Если же обе пары аллельных генов расположены в одной паре хромосом, то при образовании половых клеток гены этих аллелей не смогут свободно комбинироваться. В этом случае наблюдается сцепленное наследование. ПОЛНОЕ СЦЕПЛЕНИЕ Т. Морган скрещивал черных длиннокрылых самок с серыми с зачаточными крыльями самцами. У дрозофилы серая окраска тела доминирует над черной, длиннокрылость — над зачаточными крыльями. Обозначим ген серой окраски тела В, аллельный ему ген черной окраски тела Ь\ ген длиннокрылости V, аллельный ему ген зачаточных крыльев v. Обе пары этих генов нахо- 52 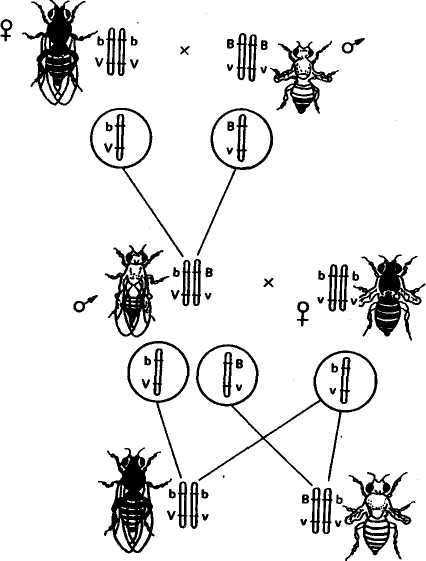 ГАМЕТЫ ГАМЕТЫ дятся в одной и той же второй паре хромосом. По обеим парам признаков родительские формы были гомозиготны: самка по рецессивному признаку черного тела (bb) и доминантному признаку длиннокрылости (VV), самец по доминантному признаку серой окраски. (ВВ) и рецессивному признаку зачаточных крыльев (w). Гаметы родителей при редукционном делении получают у материнской формы хромосому с генами b и V, у отцовской-— с генами В и v. Все потомство первого поколения (Fi) имело серое тело и длинные крылья (рис. 13) и было гетерозиготно по обеим парам признаков (bV/Bv). Затем из Fi были отобраны самцы, которых скрестили с гомозиготными по обоим рецессивным генам самками, черными зачаточнокрылыми (bv/bv), т. е. было проведено анализирующее скрещивание, в результате которого при независимом комбинировании признаков должны были бы получить потомство четырех фенотипов в Рис. 13. Наследование окраски тела и формы крыльев у дрозофилы при полном сцеплении: В—серая окраска; Ь — черная окраска; V— длинные крылья; v —короткие крылья 53 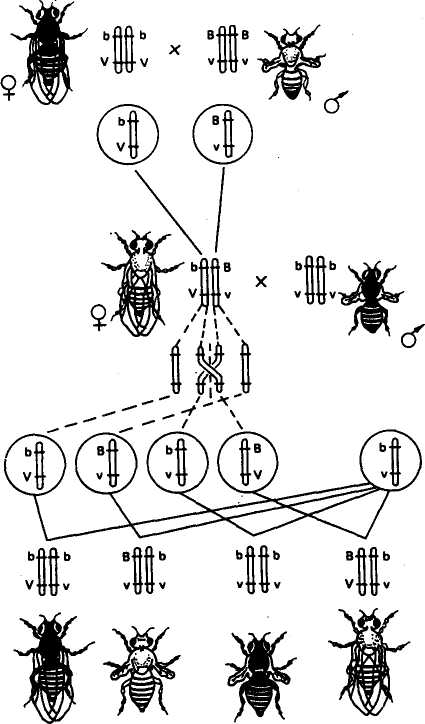 равных соотношениях: серых длиннокрылых, серых с зачаточными крыльями, черных длиннокрылых, черных с зачаточными крыльями, но были получены потомки только «двух фенотипов, похожих на исходные родительские формы: черные длиннокрылые и серые короткокрылые. В этом случае наблюдается полное сцепление признаков. Это связано с тем, что у гетерозиготного самца в одной и той же хромосоме из гомологичной пары расположены и ген черной окраски, и ген длинных крыльев, в другой — ген серой окраски и ген зачаточнокрылости. равных соотношениях: серых длиннокрылых, серых с зачаточными крыльями, черных длиннокрылых, черных с зачаточными крыльями, но были получены потомки только «двух фенотипов, похожих на исходные родительские формы: черные длиннокрылые и серые короткокрылые. В этом случае наблюдается полное сцепление признаков. Это связано с тем, что у гетерозиготного самца в одной и той же хромосоме из гомологичной пары расположены и ген черной окраски, и ген длинных крыльев, в другой — ген серой окраски и ген зачаточнокрылости.При спермиогенезе в период мейоза гомологичные хромосомы расходятся в разные половые клетки. Образуется только два сорта гамет: один с хромосомой, которая несет гены Ъ и V, другой с хромосомой, в которой расположены гены В и v. При сочетании указанных гамет с гаметами особи с рецессивными признаками и образуется потомство только двух типов. При полном сцеплении гены, расположенные в одной хромосоме, всегда передаются вместе. Полное сцепление пока установлено только у самцов дрозофилы и самок тутового шелкопряда. НЕПОЛНОЕ СЦЕПЛЕНИЕ В следующем опыте, так же как и в предыдущем, Морган скрещивал черных длиннокрылых самок с серыми зачаточно-крылыми самцами. В первом поколении получил все потомство серое длиннокрылое. Затем снова произвел анализирующее скрещивание, но из первого поколения отобрал не самца, а самку и скрестил ее с черным с зачаточными крыльями самцом (рис. 14). В этом случае появилось потомство не двух типов, как при полном сцеплении, а четырех: серое $- зачаточными крыльями, черное длиннокрылое, серое длиннокрылое и черное с зачаточными крыльями, но не в равных соотношениях, как при независимом комбинировании признаков, а со значительным преобладанием фенотипов, сходных с родительскими формами. 41,5 % мух было серых с зачаточными крыльями, как у одного исходного родителя, и 41,5 % особей черных длиннокрылых, как у другого исходного родителя. Только 17 % потомков родилось с новым сочетанием признаков: 8,5 % черных с зачаточными крыльями и 8,5 % серых длиннокрылых. Таким образом, 83 % потомков имели сочетание признаков, как у исходных родительских форм, но появились особи и с новым сочетанием признаков. Следовательно, сцепление является неполным. / Встал вопрос: почему появились особи с новой комбинацией родительских признаков? Для объяснения этого явления Морган использовал и развил теорию хиазмотипии бельгийского цитолога Янсенса. В 1909 г. Янсенс наблюдал, что при спермиогенезе у саламандры в профазе мейоза гомологичные хромосомы конъ-югируют, а затем, при начале расхождения, образуют фигуры в 54 МЕЙОЗ СТАДИЯ ЧЕТЫРЕХ ХРОМАТИД ГАМЕТЫ ГАМЕТЫ ЧЕРНЫЕ СЕРЫЕ С ЧЕРНЫЕ С ДЛИННО- ЗАЧАТОЧНЫМИ ЗАЧАТОЧНЫМИ КРЫЛЫЕ КРЫЛЬЯМИ КРЫЛЬЯМИ СЕРЫЕ ДЛИННОКРЫЛЫЕ 8,5% 41.5% 8,5% 41 .5% 17% 83% Рис. 14. Наследование окраски тела и формы крыльев у дрозофилы при неполном сцеплении: В — серая окраска; Ь — черная окраска; V— длинные крылья; v — короткие крылья виде греческой буквы «хи» (х), откуда это явление и получило название хиазмотипии, а фигуры перекреста хромосом — название хиазм. Морган на основании этих наблюдений Янсенса высказал гипотезу о том, что при образовании хиазм гомологичные 55 хромосомы обмениваются участками. Если сцепленные гены лежат в одной хромосоме и у гетерозигот при образовании гамет происходит рекомбинация этих генов, значит, гомологичные хромосомы во время мейоза обменялись своими частями. Обмен гомологичных хромосом своими частями называется перекрестом или крос-синговером (английское слово crossingover означает образование перекреста). Особей с новыми сочетаниями признаков, образовавшимися в результате кроссинговера, называют кроссоверами. Вернемся к рисунку 14. У самки Fi, гетерозиготной по обеим парам признаков, в одной из гомологичных хромосом расположены гены Ь и V, в другой — аллельные им гены В и v. В профазе редукционного деления, когда две гомологичные хромосомы соединились в один бивалент, каждая из хромосом удвоена и состоит из двух хроматид. Всего будет 4 хроматиды- Между двумя хроматидами гомологичных хромосом и происходит обмен их частями. В результате ген Ь, расположенный в хроматиде одной гомологичной хромосомы, может соединиться с геном v, расположенным в хроматиде другой гомологичной хромосомы, и как результат одного события образуется вторая хроматида, где соединятся гены В и V. В дальнейшем хроматиды разойдутся и образуются кроссоверные гаметы с хромосомами с новым сочетанием генов (bv и BV). Две другие хроматиды из пары гомологичных хромосом не участвуют в перекресте и сохраняют в первоначальном сочетании материнские (JbV) и отцовские (Bv) гены. Образование новых кроссоверных гамет обеспечило появление дрозофил с новым сочетанием признаков: черных с зачаточными крыльями и серых длиннокрылых. Однако большая часть потомков будет сходна с исходными родителями (черные длиннокрылые и серые коротко-крылые). Морган приходит к выводу, что количество появления новых форм зависит от частоты перекреста, которая определяется по следующей формуле: „ Число кроссоверных форм ■ 100 Частота перекреста = ^т3- — . Общее число потомков Если, например, общее число потомков 900, а новых кроссоверных форм 180, то частота перекреста будет составлять 20 %. Морган установил, что частота перекреста между определенной парой генов — относительно постоянная величина, но различная для разных пар генов. На основании этого был сделан вывод о том, что по частоте перекреста можно судить о расстояниях между генами. За единицу измерения перекреста принята его величина, равная 1 %. Иногда ее называют морганидой. Величина перекреста зависит от расстояния между изучаемыми генами. Чем больше отдалены гены друг от друга, тем чаще происходит перекрест; чем ближе они расположены, тем вероятность перекреста меньше. Установлено, что количество кроссоверных особей к общему 56 числу потомков никогда не превышает эи уь, так как. при ичсш> больших расстояниях между генами чаще происходит двойной кроссинговер и часть кроссоверных особей остается неучтенной. Их можно учесть при изучении не двух пар сцепленных признаков, а трех или четырех. В этом случае, учитывая двойные и тройные перекресты, можно точнее судить о расстояниях и частоте перекреста между генами. СОМАТИЧЕСКИЙ (МИТОТИЧЕСКИЙ) КРОССИНГОВЕР Сущность соматического кроссинговера заключается в том, что он осуществляется при митотическом делении соматических клеток главным образом эмбриональных тканей. Кроссинговер происходит между двумя несестринскими хроматидами гомологичных хромосом. У гетерозиготных особей наблюдаются отклонения в проявлении нормальных признаков. Явление соматического кроссинговера было предсказано А. С. Серебровским в 1922 г. при анализе причин появления исключительных перьев у кур. В 1936 г. соматический кроссинговер обнаружил К. Штерн у дрозофилы. Он исследовал самок серых с нормальными щетинками, но гетерозиготных (АаВЬ) по рецессивным генам желтой окраски тела (а) и опаленных щетинок (b). На теле некоторых серых с нормальными щетинками мух наблюдались двойные пятна. Половина пятна желтая с нормальными щетинками и половина серая, но с опаленными щетинками. Появление двойных пятен К. Штерн объяснил митотическим кроссинговером, в результате которого образуется часть клеток, гомозиготных по желтой окраске тела (аа), и часть, гомозиготных по опаленным щетинкам (bb). Эти клетки становятся родоначальницами при образовании участков тела с желтой окраской и нормальными щетинками и с нормальной серой окраской и опаленными щетинками. В этом случае проявляется действие рецессивных генов, оказавшихся в гомозиготном состоянии. Таким образом, осуществление кроссинговера в соматических клетках ведет к появлению мозаиков. Кроссинговер иногда происходит и на стадии размножения при образовании половых клеток, когда гонии еще имеют диплоидное число хромосом. В этом случае процент кроссоверных гамет может быть очень высоким. Частота митотического кроссинговера ниже мейотического, однако его также можно использовать для генетического картирования. Соматический кроссинговер имеет место у животных, растений и человека. Факторы, влияющие на кроссинговер. На кроссинговер могут заметно влиять условия внешней среды и генотипические факторы. Обнаружены гены, выполняющие роль запирателей кроссинговера, и гены, повышающие его частоту. В третьей хромосоме 57 дрозофилы выявлена мутация, которая прекращает процесс крос-синговера во всех парах хромосом. В качестве запирателей крос-синговера могут выступать некоторые перестройки хромосом. Чаще всего это бывает связано с инверсией (переворачиванием) того или иного участка в одной из гомологичных хромосом. На частоту кроссинговера могут влиять радиация, химические мутагены, концентрация солей, гормоны, лекарства. В большинстве случаев при воздействии этих факторов частота перекреста повышается. Нормальный перекрест хромосом может изменяться в зависимости от температуры, возраста, пола особи. Так, у тутового шелкопряда кроссинговер идет только у самцов и не бывает у самок. У дрозофилы кроссинговер наблюдается только у самок, однако оказалось, что при рентгеновском облучении можно вызвать его и у самцов. У мыши кроссинговер бывает у обоих полов, но интенсивнее у самок; у голубей — у обоих полов, но чаще у самцов. В гетерохроматических, в частности прицентромерных, районах хромосом частота перекреста снижена, и истинное расстояние между генами на этих участках может быть изменено. КАРТЫ ХРОМОСОМ После того как была установлена связь генов с хромосомами и обнаружено, что частота кроссинговера всегда вполне определенная для каждой пары генов, расположенных в одной группе сцепления, встал вопрос о пространственном расположении генов в хромосомах. На основе анализа генетических исследований Т. Морган и его ученик А. Стертевант выдвинули гипотезу линейного расположения генов в хромосоме. Изучение взаимоотношений между тремя генами при неполном сцеплении показало, что частота (процент) перекреста между первым и вторым, вторым и третьим, первым и третьим генами равна сумме или разности между ними. Так, в одной группе сцепления расположены 3 гена — А, В, С. Оказалось, что процент перекреста между генами АС равен сумме процентов перекреста между генами АВ и ВС, частота перекреста между генами АВ оказалась равной АС — ВС, а между генами ВС=АС— АВ. Приведенные данные соответствуют геометрической закономерности в расстояниях между тремя точками на прямой. На этом основании был сделан вывод: гены расположены в хромосомах в линейной последовательности на определенных расстояниях друг от друга. На основании анализа частоты кроссинговера между генами к настоящему времени для многих видов животных и растений построены карты хромосом. Картой хромосом называется план расположения генов в хромосоме. Кестл провел опыт анализирующего тригибридного скрещива- 58 ния кроликов с тройными рецессивами с целью выяснения сцепления между такими генами: сплошная окраска — С, гималайская окраска — с"; белый жир — Y, желтый жир — у; черная окраска — В, коричневая окраска — Ъ. С ДВОЙНЫМ ПЕРЕКРЕСТОМ С ПЕРЕКРЕСТОМ НА 2-м УЧАСТКЕ В 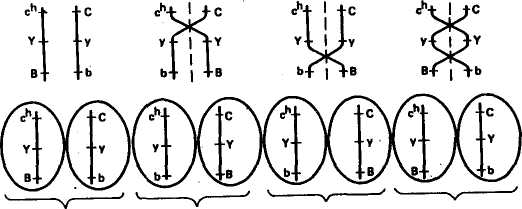 результате анализирующего скрещивания было получено 908 кроликов восьми разных фенотипов соответственно количеству разных сортов гамет (табл. 4). Численное соотношение особей разных фенотипических классов указывало на отсутствие независимого наследования по этим трем парам аллелей. Нужно было установить порядок расположения этих генов в хромосоме. Поскольку известно, что численность гамет родительских форм должна значительно превышать численность кроссоверных гамет, то можно прийти к выводу, что родительские комбинации генов были с*УВ и СуЪ (276 + 275 = 551). Они составляли от общего числа 60,7 %. Далее при анализе исходим из того, что двойных перекрестов должно быть значительно меньше, чем одинарных. Меньше всего было комбинаций сгуВ и CYb (7 + 16 = = 23) — 2,5 %. Генотипы этих кроликов отличались от родительских только тем, что Y и у поменялись местами. Так могло произойти только при двойном перекресте, и это является подтверждением того, что расположение генов было именно таким. Вычисляем частоту одиночных перекрестов. От одиночного перекреста на первом участке образовались гаметы CYB и с"уЬ (рис. 15). Случаев одиночного перекреста на первом участке было 101 (55 + 46), или 11,1 %. В результате одиночного перекреста на втором участке образовались гаметы результате анализирующего скрещивания было получено 908 кроликов восьми разных фенотипов соответственно количеству разных сортов гамет (табл. 4). Численное соотношение особей разных фенотипических классов указывало на отсутствие независимого наследования по этим трем парам аллелей. Нужно было установить порядок расположения этих генов в хромосоме. Поскольку известно, что численность гамет родительских форм должна значительно превышать численность кроссоверных гамет, то можно прийти к выводу, что родительские комбинации генов были с*УВ и СуЪ (276 + 275 = 551). Они составляли от общего числа 60,7 %. Далее при анализе исходим из того, что двойных перекрестов должно быть значительно меньше, чем одинарных. Меньше всего было комбинаций сгуВ и CYb (7 + 16 = = 23) — 2,5 %. Генотипы этих кроликов отличались от родительских только тем, что Y и у поменялись местами. Так могло произойти только при двойном перекресте, и это является подтверждением того, что расположение генов было именно таким. Вычисляем частоту одиночных перекрестов. От одиночного перекреста на первом участке образовались гаметы CYB и с"уЬ (рис. 15). Случаев одиночного перекреста на первом участке было 101 (55 + 46), или 11,1 %. В результате одиночного перекреста на втором участке образовались гаметы БЕЗ ПЕРЕКРЕСТА С ПЕРЕКРЕСТОМ НА 1-м УЧАСТКЕ Рис. 15. Типы гамет, образующихся у крольчихи, гетерозиготной по трем парам аллелей, находящихся в одной паре хромосом 59 первом участке между генами с* и у составит 13,6 % (11,1 + 2,5). На втором участке между генами у и Ъ — 28,2 % (25,7 + 2,5). Отсюда общая протяженность обоих участков, или процент перекреста между генами с* и Ъ составит 41,8 (13,6 + 28,2). 4. Фенотипические классы кроликов, полученные при анализе на сцепление трех генов Ф  енотипы енотипыСоответствующие Величина группы Распределение окраски Окраска жира Окраска тела фенотипам гаметы в разобранном выше примере кроссинговер между генами с* и у обнаружен у 13,6 % кроликов, между генами у и Ъ — у 28,2, а между генами гик учетом двойного перекреста —у 41,8 % животных. Ген Ъ не может быть расположен между генами с" и у, так как расстояние его от гена с* значительно больше, чем между генами с* и у (41,8 % против 13,6 %). Следовательно, три изученных гена расположены в хромосоме в таком порядке:
908 * Расстояние между генами с* и b можно определить и путем учета общего числа одиночных перекрестов (без включения двойных перекрестов). Оно составляет 36,8 %. Прибавив к этому удвоенный процент двойных перекрестов, т. е. 5,0 % (2,5 • 2), получим 41,8 %, что совпадает с результатами уже сделанного расчета по сумме перекрестов на каждом из участков. Теперь можно проверить, насколько фактическая величина двойного перекреста совпадает с теоретически ожидаемой. Теоретически ожидаемую величину рассчитывают путем перемножения процентов перекреста между генами на первом и втором участках, т. е. (13,6 : 100)(28,2 : 100)100 = 3,83 %. Фактически их было 2,5 %. Уменьшение числа ожидаемых двойных кроссоверов показывает, что кроссинговер на одном участке влияет на прохождение обмена на соседнем участке. Явление торможения кроссинговера на одном участке кроссинго-вером на другом получило название интерференции. Чем меньше будет расстояние, разделяющее три гена, тем больше интерференция. Принимая во внимание линейное расположение генов в хромосоме, взяв за единицу расстояния частоту кроссинговера, Морган с сотрудниками составили первую карту расположения генов в одной из хромосом дрозофилы. Затем были составлены карты других ее хромосом. Оказалось, что установленное распределение генов в хромосоме является общебиологической закономерностью. К настоящему времени составлены карты хромосом для животных и растений многих видов. Если для какого-то вида установлена группа сцепления, которая содержит три и более гена, можно составить план их расположения в хромосоме. Так, 60 28,2 13,6 Внизу цифрами указано расстояние между генами. Далее устанавливают сцепление хотя бы одного из этих генов с каким-то четвертым геном и снова проводят анализирующее скрещивание, выявляя частоту кроссинговера между вновь изучаемым геном и прежними хотя бы двумя уже изученными. На основании величины кроссинговера определяют его место в отношении к известным генам. При построении карт в хорошо изученных хромосомах указывают не расстояние между генами, а расстояние до каждого гена от нулевой точки начала хромосомы. Для лучшего усвоения материала нами везде доминантный аллель обозначается прописной буквой, рецессивный — строчной. В специальной литературе и при построении карт хромосом название (символ) гена может включать от 1 до 4 букв. Берется такое минимальное количество букв, чтобы сокращенное обозначение не совпало с любым другим обозначением генов данного вида организмов. При сокращении символа гена, как правило, берут первую букву названия признака и в случае необходимости добавляют буквы из середины или конца слова либо из других слов, входящих в название. Для обозначения генов рекомендуется использовать прилагательные или существительные латинского или английского языка. Например, гены курицы обозначают: Barring — В (оперение полосатое); Crest — О (черепно-мозговая грыжа); Prenatal — pn (гибель на поздней стадии инкубации). После построения генетических карт встал вопрос о том, отвечает ли расположение генов в хромосоме, построенное на основании частоты кроссинговера, истинному расположению. С этой целью генетические карты нужно было сравнить с цитологическими. В 30-х годах нашего столетия Пайнтер открыл в слюнных железах дрозофилы гигантские хромосомы, строение которых можно было изучать под микроскопом. Хромосомы эти имеют характерный для них поперечный рисунок в виде дисков разной толщины, которые окрашиваются кармином. Каждая хромосома по длине имеет специфические рисунки дисков, что позволяет отличать разные ее участки друг от друга. Появилась 61 |