В. Н. Сайтаниди Рецензент членкорреспондент расхн в. Ф. Красота Петухов В. Л. и др. П31 Ветеринарная генетика В. Л. Петухов, А. И. Жигачев, Г. А. Назарова. 2е изд., перераб и доп. М. Колос, 1996. 384 с ил. Учебники
 Скачать 5.3 Mb. Скачать 5.3 Mb.
|

уу черные |
 иже приводится решетка, предложенная Р. Пеннетом, где вверху по горизонтали и слева по вертикали помещены гаметы родителей, а в центральной части — генотипы потомков F2-
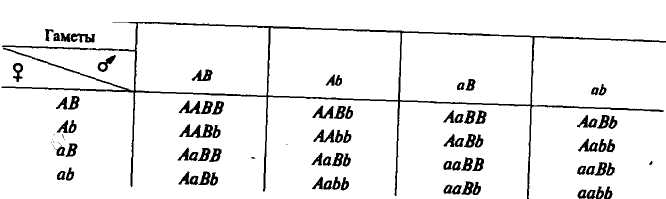
иже приводится решетка, предложенная Р. Пеннетом, где вверху по горизонтали и слева по вертикали помещены гаметы родителей, а в центральной части — генотипы потомков F2-| Расщепление | | | По одной паре аллеле{ | i |
| по генотипу | | АА | "1 2Аа | аа |
| По другой паре | | | | |
| аллелей | | | | |
| | ВВ | 1ААВВ | 2ЛаВВ | laaBB |
| | 2ВЬ | 2ЛАМ | 4АаВЬ | 2ааВЪ |
| | ЬЬ | lAAbb | 2Aabb | laabb |
Примечание. В центральной части таблицы показано расщепление по генотипу одновременно по двум парам признаков. Оно имеет такой вид: \AABB + + lAABb + \AAbb + lAaBB + ААаВЬ + TAabb + UaBB + 2aaBb + laabb.
Мендель показал, что независимое наследование признаков обусловлено независимым наследованием разных пар аллелей. В основе независимого наследования разных пар аллелей (наследственных задатков) лежит независимое комбинирование хромосом при образовании гамет.
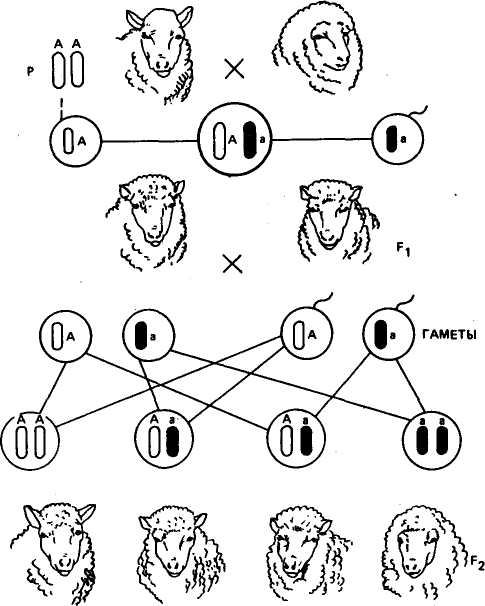
Правильность своих выводов о независимом комбинировании генов и признаков Мендель подтвердил путем проведения анализирующего скрещивания. Он скрестил гибридные растения Fi с круглыми желтыми семенами (AaBb) с отцовским растением, гомозиготным по рецессивным признакам морщинистой формы семян и зеленой их окраски (aabb). У гибрида Fi при независимом комбинировании генов равновероятно образование четырех сортов гамет (АВ, Ab, aB, ab), у отцовского растения образуются гаметы только одного сорта (ab). Наследование идет по следующей схеме:
р
 О АаВь крутые желтые х О ааЬЬ морщинистые зеленые
О АаВь крутые желтые х О ааЬЬ морщинистые зеленыеАа,
aabb
Гаметы
39
ГАМЕТЫ
Fl ? JO.
Было получено потомство четырех фенотипов: круглые желтые (АаВЬ), круглые зеленые (АаЬЬ), морщинистые желтые (ааВЬ), морщинистые зеленые {aabb). Потомков 'каждого типа было получено одинаковое число — по 25 %. Так как у отцовского растения половые клетки были одинаковые — ab, то равное число особей с каждым фенотипом является результатом того, что гибриды Fi (АаВЬ) образовали половые клетки каждого типа (AB, Ab, аВ, ab) в равном количестве. Это возможно только при независимом комбинировании генов.
На основании опытов по дигибридному скрещиванию был установлен закон независимого наследования признаков (называют также — независимого наследования аллелей разных генов).
Закон независимого наследования признаков состоит в том, что во втором поколении каждая пара аллельных генов и признаков, определяемых ими, ведет себя независимо от других пар аллельных генов и признаков. При этом возникают всевозможные сочетания в определенных числовых соотношениях по фенотипу и генотипу. При дигибридном скрещивании, при полном доминировании, соотношение по фенотипу будет 9:3:3:1, при тригибридном скрещивании будет свое определенное соотношение и т. д.
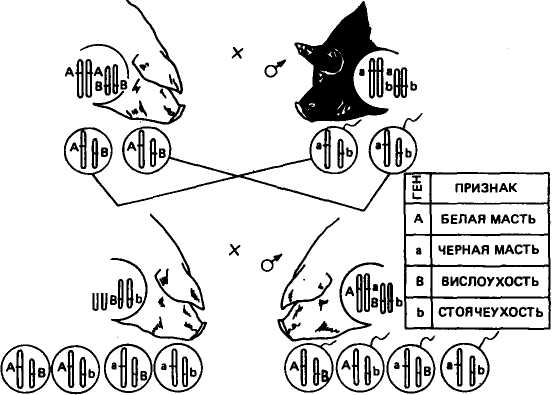
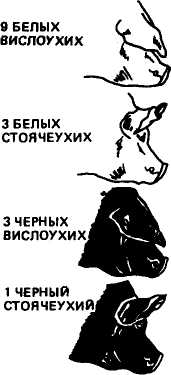
Разберем пример дигибридного скрещивания применительно к животным. Скрестим свиноматку породы ландрас белую с висячими ушами с хряком беркширской породы черным со стоячими ушами. Одна пара признаков характеризует масть (белая или черная), другая пара — форму ушей (висячие или стоячие). Ген доминантного признака белой масти обозначим прописной буквой А, а аллельный ген рецессивной черной масти — строчной буквой а. Ген доминирующих висячих ушей обозначим буквой В, ген стоячих ушей — Ь. Допустим, что аллельные гены по указанным парам признаков находятся в разных парах хромосом' (рис. 10). Оба родителя гомозиготны: мать по доминантным признакам белой масти и висячих ушей (ААВВ), отец по рецессивным признакам черной масти и стоячих ушей (aabb). В период образования половых клеток при мейозе из каждой пары гомологичных хромосом в гамету придет только одна. Поскольку родители гомозиготны, то у каждого из них будут все гаметы одного типа: у свиноматки АВ, у хряка ab.
В первом поколении в соответствии с первым законом Менделя все свиньи по фенотипу будут одинаковыми: белыми, с висячими ушами, по генотипу дигетерозиготны, т. е. гетерозиготны по обеим парам признаков (АаВЬ).
В гибридах F2, которые получают путем спаривания животных Fi между собой, наблюдается расщепление. Предположим, что одна пара аллельных генов А и а находится в более длинных гомологичных хромосомах, другая пара В и b — в более коротких. В результате мейоза из каждой пары гомологичных хромосом в каждую половую клетку пойдет только одна. Аллели А и а
40
| | АВ | АЬ | аВ | аЬ |
| АВ | ААВВ БЕЛЫЙ ВИСЛ. | ААВЬ БЕЛЫЙ ВИСЛ. | АаВВ БЕЛЫЙ ВИСЛ. | АаВЬ БЕЛЫЙ ВИСЛ. |
| АЬ | ААВЬ БЕЛЫЙ ВИСЛ. | ААЬЬ БЕЛЫЙ СТОЯЧЕУХ. | АаВЬ БЕЛЫЙ ВИСЛ. | АаЬЬ БЕЛЫЙ СТОЯЧЕУХ. |
| ав | АаВВ БЕЛЫЙ ВИСЛ. | АаВЬ БЕЛЫЙ ВИСЛ. | ааВВ ЧЕРНЫЙ ВИСЛ. | ааВЬ ЧЕРНЫЙ ВИСЛ. |
| аЬ | АаВЬ БЕЛЫЙ ВИСЛ. | АаЬЬ БЕЛЫЙ СТОЯЧЕУХ | ааВЬ ЧЕРНЫЙ ВИСЛ. | ааЬЬ ЧЕРНЫЙ СТОЯЧЕУХ. |
Рис. 10. Схема скрещивания белой с висячими ушами свиноматки с черным со стоячими ушами хряком
разойдутся, то же самое произойдет и с аллелями В и Ь. Расхождение в гаметы хромосом из каждой пары происходит независимо, поэтому аллель А может уйти с равной вероятностью как в те гаметы, куда ушел аллель В, так и в те, куда ушел аллель Ь. Аллель а также с равной вероятностью может попасть в гаметы
41
как с аллелем В, так и с аллелем Ъ. В результате как хряки, так и свиноматки из Fi образуют по четыре сорта гамет: АВ, АЬ, аВ, аЬ в равном количестве.
Каждый из спермиев может оплодотворить любую из яйцеклеток с одинаковой вероятностью. Получается 16 возможных сочетаний гамет отца и матери. Результаты этой случайной встречи гамет хорошо видны при использовании решетки Пен-нета. В верхней горизонтальной строке как заголовки записаны типы гамет одного родителя, а слева вертикально, как заголовки строчек, расположены типы гамет другого родителя. В каждый квадрат на пересечении столбца и строчки записаны генотип и фенотип потомка, определяемые исходя из сочетаний гамет, стоящих в заголовках. В заголовках располагают сначала гаметы с доминирующими генами, затем с доминирующим и рецессивным и в конце с рецессивными. При записи генотипа каждого потомка сначала располагают гены одной аллельной пары, затем другой, рядом указывают фенотип. Затем подсчитывают особей с разными фенотипами и генотипами.
Из данных решетки видно, что в F2 среди особей с разным фенотипом наблюдается следующее количественное соотношение: 9 частей белых с висячими ушами; 3 части белых со стоячими ушами; 3 части черных с висячими ушами; 1 часть черных со стоячими ушами. Рассматривая каждую пару признаков отдельно, находим, что на 12 белых свиней приходится 4 черные (отношение 3:1) и, с другой стороны, на 12 свиней с висячими ушами — 4 со стоячими (отношение 3:1).
Таким образом, каждая пара признаков при наследовании ведет себя независимо от другой пары, и только в результате их свободного комбинирования наблюдается характерное для ди-гибридного расщепления соотношение фенотипов в F2 — 9:3:3:1, которое можно рассматривать как результат сочетания двух моногибридных расщеплений (3:1 и 3:1).
Расщепление по генотипу во втором поколении точно такое же, как установил Мендель на растениях гороха.
ПОЛИГИБРИДНОЕ СКРЕЩИВАНИЕ
Положение о независимом наследовании разных пар аллелей и признаков было подтверждено Менделем при изучении наследования трех пар признаков у гороха. Он скрещивал сорт растения с круглыми семенами (А), желтыми семядолями (В) и серо-коричневой кожурой семян (С) с сортом, форма семян которого морщинистая (а), семядоли зеленые (Ь), семенная кожура белая (с). Материнское растение имело генотип ААВВСС, отцовское — ааЪЪсс. Поскольку родители гомозиготны по всем трем парам признаков, у них образуется по одному типу гамет: ABC и аЪс. Гибриды Fi будут иметь генотип АаВЬСс (тригетерозигота). При
42
тригетерозиготности три пары разных аллелей находятся в трех разных парах гомологичных хромосом. В результате независимого сочетания хромосом (значит, и аллелей) из разных пар у гибрида Fi образуется восемь сортов гамет: ABC, ABc, АЪС, аВС, Abe, aBc, abc, аЪс. При самоопылении в результате случайного сочетания гамет в F2 получается 64 комбинации, включающие 8 фенотипов.
Мендель установил, что расщепление по фенотипу при три-гибридном скрещивании представляет собой сочетание трех независимых моногибридных расщеплений. Чем больше признаков, по которым отличаются взятые для скрещивания особи, тем сложнее расщепление и сильнее возрастает комбинативная изменчивость. Число возможных комбинаций гамет и число классов по фенотипу можно определить, пользуясь таблицей 3.
3. Количество фенотипов и генотипов в F2 при скрещивании родителей,
отличающихся разным числом пар признаков
(при полном доминировании)
Число возможных комбинаций
гамет
Ч
 исло классов в F2
исло классов в F2Число гамет,
образующихся
в F2
Число пар признаков
Пропорция
по фенотипу
по генотипу
полных рецессивов в
F2
| 1 | 2'=2 | 4'=4 | 2'=2 | з'=з | 1/4 |
| 2 | 22=4 | 42=1б | 22=4 | 32=9 | 1/16 |
| 3 | 23=8 | 43=64 | 23=8 | 33=27 | 1/64 |
| 4 | 24=16 | 44=256 | 24=16 | 34=81 | 1/256 |
| л | 2" | 4я | 2я | 3я | 1/4" |
Для того чтобы понять, почему в пределах популяции каждого вида животных наблюдается такое большое разнообразие в типе телосложения, размерах, продуктивности и т. д., можно произвести простые расчеты при помощи формулы 2й. Цифра 2 показывает, что набор хромосом диплоидный, я — гаплоидное число хромосом у определенного вида животных. Если отец и мать гетерозиготны только по одной какой-то паре аллельных генов, расположенных в каждой паре хромосом, то при полном доминировании каждого из признаков число определяемых этими аллелями возможных различных фенотипов у их потомков будет: у крупного рогатого скота 230, или более миллиарда, у свиней 219, или более 500 тыс., и т. д. Но животные, очевидно, могут отличаться друг от друга и по большему числу пар аллелей. Поэтому потенциальные возможности комбинативной изменчивости огромны, и становится понятным, почему в природе не встречается абсолютно похожих особей, за исключением однояйцевых близнецов.
Огромной заслугой Менделя является то, что в процессе своей работы он не только установил закономерности наследова-
43
СТРУЧКОВИДНЫЙ ггСС
РОЗОВИДНЫЙ RRcc
©
ГАМЕТЫ
ГГ.."'
ОРЕХОВИДНЫЙ RrCc
ГАМЕТЫ F,
ОРЕХОВИДНЫЙ RrCc
ОРЕХОВИДНЫЙ RRCc
ОРЕХОВИДНЫЙ RrCC
розовидный
Rrcc
РОЗОВИДНЫЙ RRcc
ОРЕХОВИДНЫЙ RRCc
ОРЕХОВИДНЫЙ RrCc
СТРУЧКОВИДНЫЙ ггСс
ОРЕХОВИДНЫЙ RrCC
ОРЕХОВИДНЫЙ RrCc
СТРУЧКОВИДНЫР ггСС
РОЗОВИДНЫЙ Rrcc
ОРЕХОВИДНЫЙ RrCc
ЛИСТОВИДНЫЙ
СТРУЧКОВИДНЫМ гСс
 ния признаков, но и открыл основные принципы (законы) наследственности:
ния признаков, но и открыл основные принципы (законы) наследственности:
дискретной (генной) наследственной детерминации признаков.
Этот принцип лежит в основе теории гена;
относительного постоянства наследственной единицы (гена);
аллелъного состояния гена (доминантность и рецессивность).
\1
ВЗАИМОДЕЙСТВИЕ НЕАЛЛЕЛЬНЫХ ГЕНОВ
Иногда на формирование признака влияют две или несколько пар неаллельных генов. Проявление признака в этом случае зависит от характера их взаимодействия в процессе развития организма. В первом поколении появляется новый признак, которого не было у исходных родительских форм, и соотношение фенотипов во втором поколении будет иным. Рассмотрим несколько примеров различных типов взаимодействия генов.
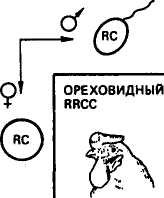
Новообразование. Новообразованием называется такой тип взаимодействия генов, когда при их сочетании в одном организме развивается совершенно новая форма признака. Известно,, что у кур гены розовидного и стручковидного гребня не являются аллельными. И стручковидный, и розовидный гребень доминирует над листовидным. При скрещивании кур породы виан* дот, имеющих розовидный гребень (RRcc), с петухами породы брама со стручковидным гребнем (rrCQ у потомков первого поколения (RrCc) в результате взаимодействия двух доминантных генов R и С развивается новая форма гребня — ореховидная (рис. 11). Скрещивание потомков Fi между собой ведет к получению в F2 четырех разных фенотипов в соотношении: 9 — с генами R и С с гребнем ореховидной формы, 3Rcc — с гребнем розовидной формы, ЪггС— со стручковидным гребнем и \rrcc <— с листовидным. Расщепление по фенотипу 9:3:3:1. В этом случае взаимодействие неаллельных генов R и С обусловливает образование новой формы гребня, в то время как каждый из этих генов в отдельности проявляет свой собственный эффект. Особь с листовидным гребнем является двойным рецессивом.
Комплементарное взаимодействие генов. В том случае, когда признак образуется при наличии двух доминантных неаллельных генов, каждый из которых не имеет самостоятельного фенотипи-ческого выражения, гены обозначают как комплементарные. Например, при скрещивании белых минорок с белыми шелковистыми курами первое поколение получается окрашенным. Для развития окраски необходимо, чтобы в организме синтезировались тирозин — предшественник меланина и фермент тирозингидрок-силаза, без которого пигмент не образуется. Обычно способность синтезировать какое-либо вещество доминирует над неспособностью к его образованию. Белые минорки имеют генотип ССоо. Они способны синтезировать тирозин, необходимый для образования
44
©
Рис. 11. Наследование формы гребня у кур при взаимодействии двух пар генов
(новообразование)
пигмента, но не способны синтезировать фермент тирозингид-роксилазу, превращающий это вещество в пигмент. Белые шелковистые куры имеют генотип ссОО. Они не способны синтезировать тирозин, но обладают способностью синтезировать фермент. При спаривании таких кур между собой (ССоо х ссОО) Fi (СсОо) получается окрашенным. В этом случае произошло образование
45
пигмента в результате включения в генотип птиц Fi обоих доминантных генов — С (обусловливающего синтез тирозина) и О (обусловливающего синтез фермента). Во втором поколении окрашенных птиц ожидается 9 частей (СО..), а белых — 7 частей (С.оо—3, ccO..—З и ссоо—1).
Белые Белые
минорки шелковистые
Р
Гаметы
окрашенные
| | СО | Со | сО | со |
| со | ССОО окраш. | ССОо окраш. | СсОО окраш. | СсОо окраш. |
| Со | ССОо окраш. | ССоо | СсОо окраш. | Ссоо Ш1 |
| сО | СсОО окраш. | СсОо окраш. | ссОО [бел\ | ссОо \Ш\ |
| со | СсОо окраш. | Ссоо | ссОо | ССОО |
Комплементарным взаимодействием генов обусловлен, очевидно, особый тип паралича задних конечностей у помесных собак, полученных от скрещивания датского дога с сенбернаром. Генетический анализ, проведенный Стокардом, показал, что при чистопородном разведении как у датских догов, так и у сенбернаров паралич не наблюдается. В то же время из 57 помесей Fi, полученных от реципрокных скрещиваний указанных пород и доживших до 3-месячного возраста, только 3 или 4 не имели такого дефекта. Среди помесей F2 из 66 щенков, доживших до 3 мес, была парализована почти треть. Болезнь проявляется внезапно в возрасте около 3 мес. Тяжесть болезни может быть различной: от слабой парализованности до полной утраты способности к самостоятельному передвижению. Аналогичное заболевание встречается у некоторых помесных собак-ищеек.
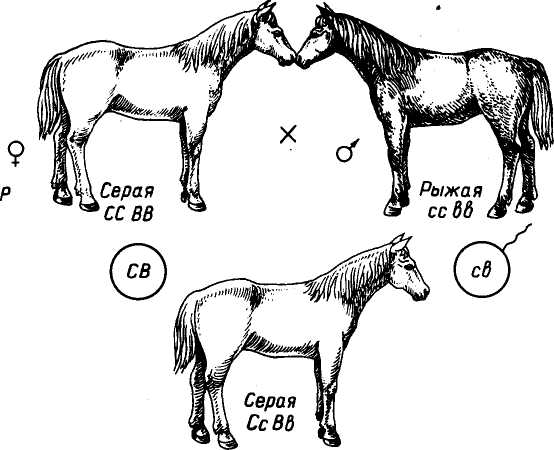
Эпистаз. При этом типе взаимодействия доминантный ген одной пары аллелей, например ген С, подавляет действие другого неаллельного доминантного гена В. При генотипе ССВВ проявляются признаки, обусловленные геном С. Ген, подавляющий развитие другого признака, называется эпистатичным, а подавляемый — гипостатичным. Например, у лошади серая доминирующая масть, связанная с ранним поседением, перекрывает все другие масти. При скрещивании серой лошади генотипа ССВВ с
46
масти. При скрещивании серой лошади генотипа ССВВ с рыжей генотипа ссЪЪ в Fi все потомки будут серыми с генотипом СсВЪ. При скрещивании Fi между собой в F2 наблюдается расщепление по фенотипу: 12 серых, 3 вороные и 1 рыжая. Аллель серой масти (С) перекрывает действие других независимых генов окраски. Все лошади, имеющие в генотипе аллель С, будут серыми. Если аллель С отсутствует, при наличии в генотипе аллеля В лошадь будет вороной (ссВВ, ссВЬ), и лошадь с генотипом ccbb, двойным рецессивом будет рыжей окраски (рис. 12).
Полимерия. При полимерии, или полимерном (полигенном) наследовании, на один и тот же признак влияют несколько разных, но сходно действующих неаллельных генов. Каждый из них усиливает развитие признака. Такие однозначно действующие гены называются аддитивными. Впервые этот тип взаимодействия генов установлен Нильсоном-Эле при изучении наследования окраски чешуи овса и зерен пшеницы.
Рассмотрим пример наследования окраски зерен пшеницы при взаимодействии двух пар полимерных генов. Различают две основные окраски зерен: красную и белую. Полимерные гены, действующие на один и тот же признак, обозначают одинаковой буквой. Разные аллельные пары обозначают цифрами внизу букв. Исходя из этого, генотип пшеницы с темно-красным зерном будет А1А1А2А2, с белым зерном — ата&г- У первого родителя образуются гаметы А1А2, у второго — в/д* В результате потомки Fi будут иметь генотип Aiayi^ (двойная гетерозигота) и промежуточную окраску зерен — светло-красную, так как имеется два доминантных гена (Ai и Аг), влияющих на проявление признака. Потомки первого поколения образуют по четыре сорта гамет {А1А2, Am, (I1A2, 0102) и при спаривании между собой дадут F2, в котором расщепление по фенотипу и генотипу будет таким: из 16 частей 1 часть темно-красных (А1А1А2А2), четыре красных QA1A1A2CI2, lAiatAiAi), 6 светло-красных (4AiOiA2O2, \AiA1a2a2, ХаыААг), 4 бледно-красных (2Aiaia2O2, 20101X202) и 1 часть белых (ауД/ДгДг). В этом легко убедиться, составив решетку Пеннета. Как видим, степень развития окраски зависит от количества доминантных генов, влияющих на формирование этого признака. При отсутствии доминантных генов окраска зерна пшеницы белая.
Полимерный тип взаимодействия генов имеет большое значение для понимания наследования количественных признаков. Эти признаки не обладают фенотипической дискретностью, и их невозможно распределить по четким фенотипическим классам. Их оценивают с помощью количественных методов учета. К количественным относятся признаки, характеризующие продуктивность животных: удой за лактацию, масса животного, настриг шерсти, масса яйца. В некоторых случаях полигенно наследуется резистентность к неблагоприятным условиям внешней среды.
47
Св
сВ
СВ

СВ
Гаметы
сВ
Серая ССВв
Серая СсВВ
ссвв
С

 ерая СсВВ
ерая СсВВСв
Серая СсВВ
'■■; у.£л •. |
Серая
Серая Сс88
С
 ерая ССВВ
ерая ССВВ■!■:,..
СВ
.

 -.-ж
-.-жСерая СсВВ

вороная ее ев
Серая Ссвв
сВ
Рыжая се 88
Вороная се Вв
Р
 ис. 12. Эпистаз при наследовании серой, вороной и рыжей масти у лошадей
ис. 12. Эпистаз при наследовании серой, вороной и рыжей масти у лошадейВсе эти признаки формируются под влиянием многих генов, каждый из которых усиливает развитие признака.
Гены-модификаторы. Гены, не проявляющие собственного действия, но усиливающие или ослабляющие эффект действия других генов, называются генами-модификаторами. Гены-модификаторы играют, по-видимому, определенную роль в формировании у животных резистентное™ к инфекционным болезням. Например, скот герефордской породы имеет белую голову, и при пастбищном содержании в условиях сильной солнечной инсоляции животные с непигментированными и слабопигментирован-ными веками болеют раком глаз. При усилении пигментации век частота заболевания уменьшается, а при интенсивной пигментации в тех же условиях болезнь не возникает. Оказалось, что интенсивность пигментации кожи вокруг глаз у белоголовых животных наследственна. Это говорит о существовании генов — модификаторов основного гена, обусловливающего белую окраску головы. Таким образом, путем селекции можно избавиться от заболевания глаз раком.
Экспрессивность и пенетрантность. Под экспрессивностью понимают степень выраженности определенного признака. Внешняя среда и гены-модификаторы могут изменить экспрессию гена, т. е. выражение признака. Изменчивость проявления му-тантного гена у разных особей — довольно частое явление. Например, у потомства дрозофилы — мутантных «безглазых» мух с сильно редуцированным количеством фасеток — содержание их варьирует от почти полного отсутствия до половины нормы.
Пенетрантность гена — это доля особей, у которых проявляется ожидаемый фенотип. При полной пенетрантности (100 %) мутантный ген проявляет свое действие у каждой особи. При неполной пенетрантности (меньше 100 %) ген проявляется фе-нотипически не у всех особей. Экспрессивность и пенетрантность гена в значительной степени зависят, по-видимому, от влияния генов-модификаторов и условий развития особей.
Плейотропия. Это влияние одного гена на развитие двух и более признаков (множественное действие гена). Так, Д. К. Беляев и А. И. Железнова установили, что у норок большинство мутаций, сопровождающихся изменением окраски волосяного покрова, рецессивно и в силу плейотропии при этом снижаются плодовитость и жизнеспособность животных.
Явление плейотропии объясняется тем, что гены плейотроп-ного действия контролируют синтез ферментов, которые участвуют в многочисленных обменных процессах в клетке и в организме в целом и тем самым одновременно влияют на проявление и развитие других признаков.
На основании рассмотренного действия генов-модификаторов, взаимодействия генов и плейотропного действия генов можно видеть, что формирование признака — очень сложное яв-
49
ление, в котором участвует не один ген, а в определенной степени весь генотип особи. Влияние в целом генотипа на развитие признака привело к формированию понятий «генотипическая среда» и «генный баланс». Под генотипической средой понимают комплекс генов организма, в котором происходит действие изучаемого гена. Генный баланс — соотношение и взаимоотношение между собой всех генов, влияющих в той или иной степени на развитие признака.
Контрольные вопросы. 1. В чем состоят особенности гибридологического метода Менделя? 2. Что означают термины «фенотип», «генотип», «аллели», «гомо-зиготность» и «гетерозиготность», «доминантность» и «рецессивность»? 3. В чем сущность законов единообразия и расщепления? 4. Что такое правило чистоты гамет? 5. С какой целью проводится анализирующее скрещивание? 6. Какое скрещивание называется дигибридным? 7. В чем заключается закон независимого наследования признаков? 8. Что такое летальные гены и как они наследуются? 9. Какое расщепление по фенотипу наблюдается при разных типах взаимодействия генов? 10. Что означают понятия «гены-модификаторы», «экспрессивность» и «пенетрантнофгь», «плейотропное действие гена»?