|
В. Н. Сайтаниди Рецензент членкорреспондент расхн в. Ф. Красота Петухов В. Л. и др. П31 Ветеринарная генетика В. Л. Петухов, А. И. Жигачев, Г. А. Назарова. 2е изд., перераб и доп. М. Колос, 1996. 384 с ил. Учебники
 ББК 45.3я73 ББК 45.3я73
П31 УДК 619:636.082(075.8)
Глава 1 ПРЕДМЕТ, МЕТОДЫ И ЗНАЧЕНИЕ ГЕНЕТИКИ
Редактор В. Н. Сайтаниди
Рецензент член-корреспондент РАСХН В. Ф. Красота
 Петухов В. Л. и др. Петухов В. Л. и др.
П31 Ветеринарная генетика /В. Л. Петухов, А. И. Жигачев, Г. А. Назарова. — 2-е изд., перераб. и доп. — М.: Колос, 1996. — 384 с: ил. — (Учебники и учеб. пособия для студентов высш. учеб. заведений). ISBN 5-10-002498-4.
Рассматриваются цитологические основы наследственности, закономерности наследования признаков при половом размножении, молекулярные основы наследственности, биотехнология, мутационная изменчивость, методы изучения изменчивости, генетика онтогенеза, иммунитета, биохимический полиморфизм, генетические аномалии, болезни с наследственной предрасположенностью и методы селекции на резистентность к болезням. Во второе издание (первое вышло в 198S г.) вошли вопросы биотехнологии.
Для студентов вузов по специальности «Ветеринария».
ББК 45.3я73
Без объявления
П-
ISBN 5-10-002498-4
© ВО «Агропромиздат», 1985
© Издательство «Колос», 1996, с дополнениями
3706000000 - 049 035(01) — 96
Предмет генетики. Генетика (от греч. genesis — происхождение) — наука о наследственности и изменчивости организмов. Термин «генетика» предложил в 1906 г. У. Бэтсон. Наследственность — свойство живых существ обеспечивать материальную и функциональную преемственность между поколениями, а также обусловливать специфический характер индивидуального развития в определенных условиях внешней среды. Наследственность — это воспроизведение жизни (Н. П. Дубинин). Изменчивость — это возникновение различий между организмами по ряду признаков и свойств.
Наследственность, изменчивость и отбор —основа эволюции. Благодаря им возникло огромное разнообразие живых существ йа Земле. Мутации поставляют первичный материал для эволюции. В результате отбора сохраняются положительные признаки и свойства, которые благодаря наследственности передаются из поколения в поколение. Знание закономерностей наследственности и изменчивости способствует более быстрому созданию новых пород животных, сортов растений и штаммов микроорганизмов.
С. М. Гершензон выделяет четыре основные теоретические проблемы, изучаемые генетикой:
хранения генетической информации (где и каким образом
закодирована генетическая информация);
передачи генетической информации от клетки к клетке, от
поколения к поколению;
реализации генетической информации в процессе онтоге
неза;
4) изменения генетической информации в процессе мутаций.
Бурное развитие генетики связано с тем, что она открывает
•оаможность познания явлений жизни и намечает пути управле-Ивк ею. В настоящее время генетика занимает центральное место Ввдкыгогии. Наблюдается все более тесная интеграция генетики, селекции, ветеринарии, биохимии и других наук. В результате Интеграции генетики и ветеринарии возникла ветеринарная генетика.
Ветеринарнаягенетика — наука, изучающая наслед-
  

ственные аномалии и болезни с наследственным предрасположением, разрабатывающая методы диагностики, генетической профилактики и селекции животных на устойчивость к болезням. Задачи ветеринарной генетики следующие:
изучение наследственных аномалий;
разработка методов выявления гетерозиготных носителей
наследственных аномалий;
3) контролирование (мониторинг) распространения вредных
генов в популяциях и их элиминация;
цитогенетический анализ животных в связи с заболевания
ми;
изучение генетики иммунитета;
изучение генетики патогенности и вирулентности микроор
ганизмов, а также взаимодействие микро- и макроорганизмов;
изучение болезней с наследственным предрасположением;
разработка методов раннего выявления (т. е. маркеров) ус-
тойчивостшр* восприимчивости организма к болезням, в том
числе при (Ксутствии инфекционного фона;
изучение влияния вредных экологических веществ на на
следственный аппарат животных;
изучение генетически детерминированных реакций живот
ных на лекарственные препараты;
создание устойчивых к болезням, с низким генетическим
грузом и приспособленных к определенным условиям среды
стад, линий, типов, пород. Последние две проблемы — предмет
изучения селекционно-ветеринарной генетики;
использование методов биотехнологии для повышения ре-
зистентности животных к болезням и т д.
V Методы генетики. Явления наследственности и изменчивости на молекулярном, клеточном, организменном и популяционном уровнях изучают, используя следующие основные методы.
Гибридологический анализ основан на использовании системы скрещивания в ряде поколений для определения характера наследования признаков и свойств. Гибридологический анализ — основной метод генетики.
Генеалогический метод заключается в использовании родословных для изучения закономерностей наследования признаков, в том числе наследственных болезней. Этот метод в первую очередь применяется при изучении наследственности человека и медленно плодящихся животных.
Цитогенетический метод служит для изучения строения хромосом, их репликации и функционирования, хромосомных перестроек и изменчивости числа хромосом. С помощью цитогенетики выявляют разные болезни и аномалии, связанные с нарушением в строении хромосом и изменением их числа.
Популяционно-статистический метод приме-

Томас Гейт Морган (1866-1945)
Грегор Иоганн Мендель (1822-1884)
няется при обработке результатов скрещиваний, изучении связи между признаками, анализе генетической структуры популяций, распространении генетических аномалий в популяциях и т. д.
И м му н о ге н етич е с к и й метод включает серологические методы, иммуноэлектрофорез и др., которые используют для изучения групп крови, белков и ферментов сыворотки крови тканей. С его помощью можно установить иммунологическую несовместимость, выявить иммунодефицита, мозаицизм близнецов и т. д.
Онтогенетический метод используют для анализа действия и проявления генов в онтогенезе при различных условиях среды. Для изучения явлений наследственности и изменчивости используют биохимический, физиологический и другие методы.
Этапы развития генетики. Датой рождения генетики принято считать 1900 г., когда Г. де Фриз, К. Корренс и Э. Чермак переоткрыли законы Г. Менделя (1865). В развитии генетики можно выделить три этапа:
первый (с 1900 по 1925 г.) — этап классической генетики. В этот период были переоткрыты и подтверждены на многих видах растений и животных законы Г. Менделя, создана хромосомная теория наследственности (Т. Г. Морган);
второй (с 1926 по 1953 г.) — этап широкого развертывания работ по искусственному мутагенезу (Г. Меллер и др.). В это время было показано сложное строение и дробимость гена, заложены основы биохимической, популяционной и эволюционной
г енетики, доказано, что молекула ДНК является носителем наследственной информации (О. Эвери и др.), были заложены основы ветеринарной генетики; енетики, доказано, что молекула ДНК является носителем наследственной информации (О. Эвери и др.), были заложены основы ветеринарной генетики;
третий (начиная с 1953 г.) -^ этап современной генетики, для которого характерны исследования явлений наследственности на молекулярном уровне. Была открыта структура молекулы ДНК (Ф. Крик, Дж. Уотсон), расшифрован генетический код (Ф. Крик, М. Ниренберг, С. Очоа, Д. Маттеи и др.), химическим путем синтезирован ген (Г. Корана).
бмномп Вавилов 7-1943)
В настоящее время успешно развивается генетическая инженерия, давшая возможность переносить гены из одного организма в другой. Значительные достижения имеются в области генетики микроорганизмов, растений.
Большой вклад в развитие генетики внесли отечественные ученые. Научные генетические школы созданы Н. К. Кольцовым, Ю. А. Филипченко, Н. И. Вавиловым, А. С. Серебров-ским. Г. А, Надсон и Г. С. Филиппов получили искусственным путем мутации. Н. И. Вавилов сформулировал закон гомологических рядов наследственной изменчивости. Г. Д. Карпеченко предложил метод преодоления бесплодия у отдаленных гибридов. А. С. Серебровский и др. показали сложное строение и дроби-мость гена. С. С. Четвериков — основатель учения о генетике популяций. Б. Л. Астауров на тутовом шелкопряде доказал возможность искусственного регулирования пола. Большой вклад в становление ветеринарной генетики в нашей стране внес академик Л. К. Эрнст. В Новосибирске создан первый НИИ ветеринарной генетики и селекции.
Значение генетики для практики. Большое значение имеют теоретические исследования по проблемам генетической инженерии в селекции растений, микроорганизмов и животных, разработке более эффективных методов и средств предупреждения болезней и лечения животных. В большой степени от успешного развития генетики зависят решение проблемы пищевых ресурсов, охрана здоровья человека и животных, борьба с наследственными болезнями, охрана окружающей среды.
Фундаментальные открытия в современной генетике реализуются в селекции растений, животных и микроорганизмов. За последние годы созданы гибриды ячменя и пшеницы, ячменя и
ржи, выведены сорта пшеницы, способные давать более 100 ц зерна с 1 га, высокомасличные сорта подсолнечника с содержанием жира в семенах до 55 %, сорт подсолнечника, масло которого по составу сходно с оливковым. Выведены фитофтороу-стойчивые и ракоустойчивые сорта картофеля, триплоидная сахарная свекла и много других сортов растений. В растениеводстве широко используется явление тотипотентнос-ти, т. е. способности любой соматической клетки дать начало растению. Разработан метод микроклонального размножения нового сорта винограда, устойчивого! к филлоксере.
Методы генетической инженерии широко применяются в биотехнологии (область научно-технического прогресса, использующая биологические процессы для промышленных целей). Методом генетической инженерии во ВНИИ генетики и селекции промышленных микроорганизмов создан промышленный штамм кишечной палочки, продуцирующий аминокислоту 1-треонин (до 30 г/л раствора), а также штамм — продуцент витамина В2 — рибофлавина. В Институте биоорганической химии создан штамм кишечной палочки, синтезирующий интерферон человека. Созданы штаммы бактерий, продуцирующие аминокислоту лизин, гормон роста человека соматотропин, бактерии, превращающие целлюлозу в сахар, и т. д. Ведутся работы по введению в пекарские дрожжи генов, кодирующих такие белки, как оваль-бумин (белок куриного яйца) и миозин (белок мышц). Получены штаммы бактерий, синтезирующие инсулин человека. Успешно разрабатываются методы микробиологического синтеза вакцин и сывороток.
В животноводстве методы генетики используют:
при выведении линий и пород животных, устойчивых к
болезням;
для уточнения происхождения животных;
3) при оценке производителей по качеству потомства;
•4) при цитогенетической аттестации производителей;
в пушном звероводстве;
для изучения влияния экологически вредных веществ на
наследственный аппарат животных и т. д.
В настоящее время генетика занимается изучением следующих основных проблем:
1) проводятся обширные исследования в области генетической инженерии с целью получения в достаточном количестве инсулина, интерферона, антибиотиков, витаминов, незаменимых аминокислот, кормовых и пищевых белков, биологических средств защиты растений и т. д.;
• 2) решается одна из стратегических задач генетики — регуля-нМя и управление действием генов в онтогенезе. Необходимо выяснить пути реализации генетической информации в признаках в процессе онтогенеза. Создание методов управления дейст-
вием генов позволит повысить продуктивность животных, резис-тентность к болезням, подавить проявление нежелательных признаков;
ставится задача разработать методы управления процессами
мутаций, что даст возможность получать нужные наследственные
изменения при создании новых штаммов микроорганизмов, сор
тов растений, линий и пород животных;
изучается проблема регуляции пола у животных. Она пока
решена в отношении регуляции пола у шелкопряда;
ведутся перспективные исследования по генокопированию
у животных, т. е. пересадка в яйцеклетку, из которой удален
собственный генетический материал, ядра, взятого из соматичес
кой клетки. Такие манипуляции уже проводят у амфибий, рыб,
мышей. Разрабатываются методы получения генетических копий
выдающихся по продуктивности и устойчивости к болезням жи
вотных;
решается проблема зашиты наследственности человека и
животных ож лутагенного действия радиации и химических мута
генов среды;
исследуются вопросы борьбы с наследственными болезня
ми у человека и животных, создания линий, пород, устойчивых
к болезням.
В учебнике изложены основы общей генетики, биотехнологии, биометрии и ветеринарной генетики.
Контрольные вопросы. 1. Что является предметом генетики? 2. Что изучает ветеринарная генетика? 3. Каковы основные методы изучения генетики? 4. Что вы знаете об этапах развит» генетики? 5. Какое значение имеет генетика для практики?
Глава 2
ЦИТОЛОГИЧЕСКИЕ ОСНОВЫ НАСЛЕДСТВЕННОСТИ
Организм животных состоит из миллиардов клеток, и все они ведут начало от двух половых клеток (гамет) — отцовского с'пер-мия и материнской яйцеклетки. При слиянии гамет — оплодотворении начинаются сложные процессы размножения, деления клеток и их структур, дифференциации тканей, формирования органов, становления видовых, породных и индивидуальных особенностей будущих телят, жеребят, поросят и т. д. Эти процессы реализуются под строгим генетическим контролем. Существуют материальные носители наследственности, которые локализованы главным образом в ядрах клеток. Важную роль в реализации генетической программы выполняют органоиды клетки: рибосомы, митохондрии (рис. 1).
Анализ генетических структур клеток, их количественной и качественной изменчивости составляет предмет исследований цитогенетики. В последнее время возникло новое направление — ветеринарная цитогенетика, основная задача которой — изучение распространения хромосомной патологии в породах животных.
РОЛЬ ЯДРА И ЦИТОПЛАЗМЫ В НАСЛЕДСТВЕННОСТИ
Ядро — основной компонент jcJieTKH^jie^yjirrHfiMreHeTH4ecKyro информацию. Оно^может^аходиться^ двух состояниях: покоя — интерфазы_ж_Дбления — митоза или мейоза. Интерфазное ядро представляет собой круглое "образование" с многочисленными глыбками белкового вещества, названного хроматином. Выделя-Ж?т два типа хроматина: гетерохроматин и эухроматин. Первый из Wax можно наблюдать в интерфазном ядре под световым, вто-!рой — только под электронным микроскопом. Гетерохроматин и §ухроматин выполняют разные функции в генетическом контро-Щ биосинтеза белков.
Детальнде_.изучение ядра_под э^ектр^нным_мЕидЕюскопом показало, что хроматин состоит из'очень "тонких нитей, получив-'ЧВТТ ""звание хромосом. Именно в них заложена основная часть -Шнетической информации индивидуума.
k^6 тельца, называемые
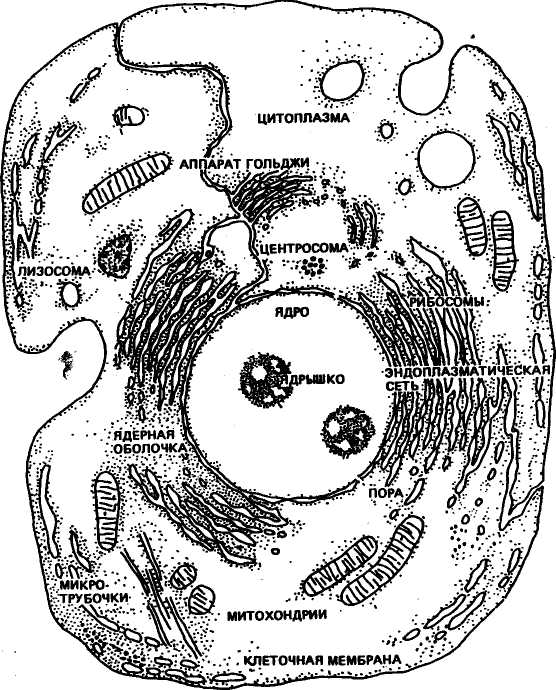
Рис. 1. Схема строения животной клетки по данным электронной
ядрышками. Количество их в зависимости от типа клеток неодинаково. По современным данным, на ядрышках осуществляется синтез рибосомной рибонуклеиновой кислоты (рРНК), а также ядерных белков (гистонов). Участки, или районы, хромосом, где происходит синтез рРНК, называют организаторами ядрышка. Например, у свиней организаторы ядрышка найдены на3=й и 10-й хромосомах. Учеными обнаружены изменчивость в ядрыш-кообразующих районах хромосом свиней и связь этого явления с отдельными болезнями, в частности с прогрессирующей атаксией и синдромом нарушения координации движений.
10
МОРФОЛОГИЧЕСКОЕ СТРОЕНИЕ ХРОМОСОМ
При микроскопическом анализе хромосом прежде всего видны различия их по форме и величине. Строение каждой хромосомы сугубо индивидуальное. Можно заметить также, что хромосомы обладают общими морфологическими признаками. Они состоят из двух нитей — хроматид, расположенных параллельно и соединенных между собой в одной точке, названной центромерой или первичной перетяжкой. На некоторых хромосомах можно видеть и вторичную перетяжку. Она является характерным признаком, позволяющим идентифицировать отдельные хромосомы в клетке. Если вторичная перетяжка расположена близко к концу хромосомы, то дистальный участок, ограниченный ею, называют спутником. Хромосомы, содержащие спутник, обозначаются как АТ-хромосомы. На некоторых из них в тело-фазе происходит образование ядрышек.
Концевые участки хромосом имеют особую структуру и называются теломерами. Теломерные районы обладают определенной полярностью, препятствующей их соединению друг с другом при разрывах или со свободными концами хромосом. Участок хрома-тиды (хромосомы) от теломеры до центромеры называют плечом хромосомы. Каждая хромосома имеет два плеча. В зависимости от соотношения длин плеч выделяют три типа хромосом: 1) мета-центрические (равноплечие); 2) субметацентрические (неравноплечие); 3) акроцентрические, у которых одно плечо очень короткое и не всегда четко различимо.
На Парижской конференции по стандартизации кариотипов вместо морфологических терминов «метацентрики» или «акро-центрики» в связи с разработкой новых методов получения «полосатых» хромосом предложена символика, в которой всем хромосомам набора присваивается ранг (порядковый номер) по порядку убывания величины и в обоих плечах каждой хромосомы (р — короткое плечо, q — длинное плечо) нумеруются участки плеч и полосы в каждом участке по направлению от центромеры. Такая система обозначений позволяет детально описывать аномалии хромосом.
Наряду с расположением центромеры, наличием вторичной перетяжки и спутника важное значение для определения отдельных хромосом имеет их длина. Для каждой хромосомы определенного набора длина ее остается относительно постоянной. Измерение хромосом необходимо для изучения их изменчивости в онтогенезе в связи с болезнями, аномалиями, нарушением воспроизводительной функции.
и Дифференциальная окраска хромосом. В последние годы для более точной идентификации хромосом применяют специальные методы обработки и окрашивания хромосом. Каждая хромосома при этом приобретает свой специфический рисунок — чередова-
11
ние светлых и темных полос, отражающих различную функциональную активность отдельных районов хромосом. Окрашенные участки — это низкоактивные в генетическом отношении гетерохроматиновые районы хромосом, а неокрашенные — сильноактивные эухроматиновые районы. Гетерохроматин, как показало дифференциальное окрашивание, существует в двух формах: 1) конститутивной — постоянно действующей в хромосоме и 2) факультативной, которая выявляется лишь в части клеточного цикла или в одной из пар хромосом.
15161718
19 20 Z1 22 23
25
Разработано несколько методов дифференциальной окраски
61, XX)
Рис. 2. G-Окраска хромосом крупного рогатого скота (2л -(по С. Г. Куликовой)
12

; Рис. 3. Комбинированная окраска хромосом крупного рогатого скота
(по С. Г. Куликовой)
хромосом: G, С, R, Q, NOR и др. (рис. 2, 3). Каждый из них имеет свое назначение. Так, полосы, окрашиваемые при С-ок-раске, идентифицируют со структурным, или конститутивным, гетерохроматином. NOR-Окраска позволяет выявить ядрышко-образующие районы хромосом. С помощью дифференциальной окраски можно не только идентифицировать отдельные хромосомы, но и, что более важно, выявить незаметные при обычной окраске поломки и перестройки хромосом; установить, какие Хромосомы в избытке или недостатке; изучить изменчивость по гетерохроматиновым районам и т. д. и связь их с морфологическими и функциональными признаками.
•Тонкое строение хромосом. Химический анализ структуры хромосом показал наличие в них двух основных компонентов:
13
дезоксирибонуклеиновой кислоты (ДНК) и белков типа гистонов и протомите (в половых клетках). Исследования точкой субмолекулярной структуры хромосом привели ученых к выводу, что каждая хроматида содержит одну нить — хромонему. Каждая хро-монема состоит из одной молекулы ДНК. Структурной основой хроматиды является тяж белковой природы. Хромонема уложена в хроматиде в форму, близкую к спирали. Доказательства этого предположения были получены, в частности, при изучении мельчайших обменных частиц сестринских хроматид, которые располагались поперек хромосомы.
КАРИОТИП И ЕГО ВИДОВЫЕ ОСОБЕННОСТИ
Анализ хромосом в клетках животных и растений разных видов позволил выявить ряд общих закономерностей, имеющих важное значение при изучении явлений наследственности и изменчивости^ Установлено, что количество хромосом в клетках разных тка^ й одного вида одинаково. Например, у домовой мыши подсчитывали число хромосом в клетках костного мозга, селезенки, лимфатических узлов, печени, почек, зобной железы, пейеровых бляшек, роговицы, эпителия кишечника; все они содержали по 40 хромосом. Форма и размеры хромосом в пределах вида также постоянны. Каждая хромосома в клетке отличается от других хромосом по внешнему строению и функциональным особенностям, т. е. индивидуальностью, которая сохраняется от одной генерации клеток к другой и передается от родителей к потомкам.
В соматических клетках хромосомы парные, а набор хромосом в них диплоидный (2я). Пары одинаковых по форме и величине хромосом называют ?$мппп:>"1'»ым" Парность хромосом возникает при слиянии (оплодотворении) мужской и женской половых клеток, которые содержат гаплоидный набор хромосом (л). Таким образом, к закономерностям строения хромосомных наборов можно отнести постоянство числа хромосом, парность, индивидуальность и непрерывность хромосом. Изучение хромосомных наборов у самцов и самок одного вида показывает, что различаются они только по одной паре хромосом. Их обозначают X (икс) и Y (игрек). Другими словами, хромосомы, по-разному представленные у двух полов и противоположно участвующие в генетическом контроле половой дифференциации и половых функций, называют половыми хромосомами или гоносомами; хромосомы, одинаковые у разных полов, — аутосомами.
При анализе наборов хромосом в клетках разных видов были выявлены различия по числу хромосом или их строению либо те и другие одновременно. Совокупность количественных и структурных особенностей диплоидного набора хромосом вида получила название кариотипа. По определению С. Г. Навашина, ка-
14
риотип — это структура — своеобразная формула вида. В карио-типе заложена генетическая информация особи, изменения которой влекут за собой изменения признаков и функций организма данной особи или ее потомства. Поэтому так важно знать особенности нормального строения хромосом, чтобы дри возможности суметь выявить изменения в кариотипе.
Для анализа хромосом важное значение имеет микрофотографирование. Оно позволяет детально изучить морфологию, подсчитать число хромосом в метафазной пластинке, измерить каждую из них. После визуальной оценки, измерений хромосомы можно вырезать и разложить по парам гомологов (идиограмма) в порядке убывающей величины (рис. 4). В таблице 1 дана характеристика кариоти-пов основных видов сельскохозяйственных животных.
1. Диплоидные наборы хромосом у сельскохозяйственных и некоторых видов домашних, прирученных и лабораторных животных
В том числе по форме том числе по форме
Аугосомы
Число хромосом (2л)
Половые хромосомы
мета-центрического и субмета-центрического типов
мета-центрического и субмета-центрического типов
Вид животного
акро-центри-ческого
типа
акро-центри-ческого
58 58
58 46 12
36 38
58 60
60 60
60 54 38
64 50
60
74
62 60 36 70 36 78 78 80 80
82
типа
XY
Европейский крупный рогатый осот (Bos taurus L.)
: крупный рогатый скот
indicus)
Y Y XY
X X
> (Сагра hircus)
24
26 10
i (Ovis aries) Домашняя свинья (Sus scrofa do-
Y X,Y
Лошадь (Equus caballus) Кавказский буйвол (Bubalus
XY XY
XY XY XY XY
12 40 26
Як (Bos grunniens)
Верблюд двугорбый (Camelus bac-
20 58 8 68
Овет (Equus asinus) %^0ison bison) ДиЯй кабан (Sus scrofa scrofa) Фверный олень (Rangifer tarandus) «noa (Felis catus)
m(Canis)
[ (Gallus gallus domesticus)
ЙКН (Meleagris gallopago)
ii*" (Anas platyrhynchus domesti-СШ)
*V« (Anser anser)
15
ас
ас эе
с
4
» t!
•<
* «о x «к
* S3
*= с:
S $
*t?f
с « € <
11
S 3
'5? 5
f % i i
S S S «о £ S
со
2 «в « « ■ t*e «
« « «в • »«в •*
с •с
в^1 t
(Л
8 .
ее I
< -
X
8
s
с
с
С
с
с с
<
*
5
митоз
В основе роста и дифференцировки органов и тканей животных лежит размножение клеток, смена одного клеточного поколения на другое. Клетки тела, или соматические клетки, разных поколении содержат одинаковое количество генетического материала, что обеспечивается особым механизмом деления, получившим название митоз. В процессе митоза выделяют две основные стадии — интерфазу и собственно митоз.
Интерфаза предшествует митозу. В ней выделяют три периода: 1) пресинтетический (Gi); 2) синтетический (S) и 3) постсинтетический (G2). В Gi-периоде в клетке происходит накопление белка, РНК и других продуктов, необходимых для образования клеточных структур и последующего деления. В течение S-периода синтезируется ДНК и происходит ауторепродукция (самоудвоение) хромосом, что приводит к возникновению второй хроматиды. В сЬ-пе-риоде продолжается синтез ДНК и белков, накапливается энергия.
Время прохождения клетками разных периодов интерфазы неодинаково. Так, по данным Л. В. Трофимовой, клетки костного мозга кур проходят периоды Gi, S, G2 и митоза соответственно за 6, 7, 8 и 23 ч. ДНК в хромосомах также синтезируется с разной скоростью. Например, у коров наибольшей" скоростью репликации характеризуется 14-я хромосома. Задержка в репликации (синтезе) ДНК на отдельных хромосомах может быть причиной возникновения патологии у животных.
Вслед за интерфазой начинается деление клетки — митоз. Выделяют четыре стадии митоза: профазу, метафазу, анафазу, тело-фазу. При изучении митоза основное внимание уделяется поведению хромосом. В профазе хромосомы представляют собой клубок длинных тонких хроматиновых нитей. К концу этой фазы митоза длина их уменьшается за счет спирализации примерно в 25 раз, наблюдается также разрушение ядрышка. Предполагают, что ве-ЩВство ядрышка участвует в образовании веретена деления. Нити веретена прикрепляются к центриолям, которые в этот период уже разделились и находятся на противоположных полюсах клет-Кй. Завершается профаза разрушением ядерной оболочки клетки.
В метафазе утолщенные спирализованные хромосомы перемещаются в экваториальную плоскость клетки. В этот момент Они имеют характерную для каждой из них форму, удобную для цитогенетического анализа.
Началом анафазы считают момент разделения удвоенных хромосом на хроматиды, которые затем расходятся к противоположным полюсам клетки.
Во время телофазы сестринские хроматиды достигают противоположных полюсов и деспирализуются. Так формируются два Дочерних ядра. Наряду с делением материнского ядра происходит деление цитоплазмы, образование оболочек клеток.
17
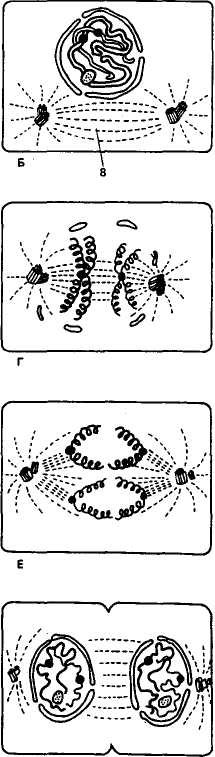
Таким образом, в процессе митоза из одной материнской клетки возникают две дочерние, содержащие такой же набор хромосом, как и у исходной клетки (рис. 5). Основное биологическое значение митоза состоит в точном распределении хромо-

Рис. 5. Схема митотического деления животной клетки (по К. Вилли и В. Детье, 1975):
А — интерфаза (стадия «покоя»); Б — ранняя профаза (центриоль разделилась); В и Г— более поздняя профаза; Д — метафаза; Е и Ж— ранняя и поздняя анафазы; 3 — телофаза (ядерная оболочка исчезла, началось деление цитоплазмы); / — хромосома; 2 — центромера; 3 — ядрышко; 4 — ядро; 5 — ядерная оболочка; 6 — центриоль; 7 — звезда; 8 — веретено
18
сом между двумя дочерними клетками; тем самым сохраняются преемственность хромосомного набора в ряду клеточных поколений и полноценность генетической информации каждой клетки, что необходимо для осуществления общих и специфических функций живого организма.
Патология митоза. При делении соматических клеток могут возникать различные нарушения, связанные с повреждением хромосом, митотического аппарата, цитоплазмы. К числу этих нарушений относятся задержка митоза в профазе, нарушения спирализации и деспирализации хромосом, раннее разделение хроматид, фрагментация и пульверизация хромосом, задержка митоза в метафазе и др. Эти нарушения возникают под действием отдельных химических веществ, радиации, вирусных инфекций. Так, при заражении свиней вирусом чумы, наблюдали пульверизацию и фрагментацию (множественные разрывы) хромосом.
ГАМЕТОГЕНЕЗ И МЕЙОЗ
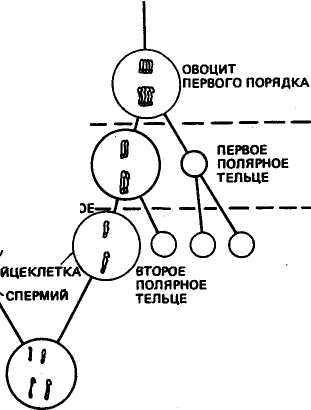
Процесс развития половых клеток носит название гаметогене-за. У самцов этот процесс называется спермиогенезом, а у самок — овогенезом(рис. 6). Половые клетки в развитии последовательно проходят следующие стадии: размножения, роста, созревания и формирования. В стадии размножения клетки интенсивно делятся митотическим путем. В стадии роста клетки накапливают питательные вещества, особенно при овогенезе.
Наиболее ответственный момент с точки зрения генетики при образовании половых клеток — мейоз — процесс редукционного и эквационного деления ооцитов и сперматоцитов, в результате которого образуются половые клетки с гаплоидным набором хромосом. Рассмотрим наиболее важные моменты поведения хОомосом в мейозе. В этом процессе выделяют две стадии деления (см. схему): 1) редукционную и 2) эквационную.
Схема мейоза
Второе эквационное деление фазы:
профаза II
метафаза II
анафаза II
телофаза II
Деление хромосом на хроматиды и образование из двух дочерних еще двух новых клеток с гаплоидным набором хромосом
редукционное деление
профаза I — лептонема
зигонема
пахинема
диплонема
диакинез метафаза I анафаза I телофаза I
Образование из одной материнской RXB с диплоидным набором хромо- ДЧ« дочерних с гаплоидным набо- удвоенных хромосом
19



ОВОГЕНЕЗ
СПЕРМАТОГЕНЕЗ
СПЕРМАТОГОНИИ В СЕМЕННИКЕ И ОВОГОНИИ В ЯИЧНИКЕ MHOFOKPAT НО ДЕЛЯТСЯ МИТОТИЧЕСКИ
ОВОГОНИИ ДАЕТ,
СПЕРМАТОГОНИИ ДАЕТ
сперматоцит /ии первого порядка!
ПЕРВОЕ
МЕЙОТИЧЕСКОЕ
ДЕЛЕНИЕ
ОВОЦИТ ВТОРОГО ПОРЯДКА ВТОРОЕ МЕЙОТИЧЕСК1 ДЕЛЕНИЕ
СПЕРМАТОЦИТ ВТОРОГО* Г« ПОРЯД1
СПЕРМАТИДЫ,
ЗИГОТА
Рис. 6. Сравнение процессов сперматогенеза и овогенеза у животных с гаплоидным числом хромосом, равным 2 (по К. Вилли и В. Детье, 1975)
Непосредственно перед мейозом клетки половых желез находятся в интерфазе.
Редукционное деление начинается с профазы I, которая, как видно из схемы, подразделяется на пять фаз. На первой стадии профазы I — лептонемы хромосомы деспирализо-ваны, они в 2—5 раз длиннее метафазных. Под электронным микроскопом можно видеть, что они состоят из двух хроматид, соединенных центромерой. На следующей стадии — зигонемы наблюдаются притяжение и слияние (конъюгация) гомологичных хромосом. Каждая пара конъюгирующих хромосом образует бивалент, а по числу хроматид — тетраду. На этой стадии происходит образование синаптонемного комплекса (СК), входящего в состав бивалента. Нарушение формирования СК между гомологичными хромосомами наблюдают у гетерозиготных носителей хромосомных аберраций. Далее, на стадии пахинемы, происходят
20
утолщение и укорочение хромосом, так что сестринские хроматиды становятся хорошо различимыми; на отдельных из них можно видеть и ядрышки.
Следующая фаза — диплонема характеризуется тем, что конъ-югярующие хромосомы начинают отталкиваться и постепенно расходятся от центромеры к концам. При этом образуются характерные фигуры, напоминающие греческую букву «хи» (х) и получившие вследствие этого название хиазмы. В точках соприкосновения гомологичных хромосом возникают разрывы. Они могут быть одинарными, двойными и более сложными. В результате разрывов образуются фрагменты хроматид, которые затем могут воссоединяться на другой хромосоме, изменяя тем самым комбинацию генетического материала в клетке.
Обмен участками между гомологичными хромосомами получил название кроссинговера.
На последней стадии профазы I — диакинезе происходит резкое укорочение хромосом, так что к концу этой стадии хроматиды остаются связанными только на концевых участках. Этим и заканчивается профаза I. Необходимо отметить, что при более детализированном изучении мейоза в профазе выделяют и другие промежуточные стадии, например пролептонему, диктионему и т. д.
На стадии метафазы I биваленты располагаются в плоскости экватора центромерами к противоположным полюсам. Силы отталкивания здесь увеличиваются.
В анафазе I начинается расхождение гомологичных хромосом к противоположным полюсам, которое носит случайный характер. Каждая из пар гомологичных хромосом имеет одинаковую вероятность распределения в одну из двух дочерних клеток.
В телофазе I хромосомы достигают полюсов клетки. Затем восстанавливаются ядерная оболочка и ядрышко, хромосомы декон-денсируются. В конце телофазы делится цитоплазма {цитокинез) и образуются две дочерние клетки с гаплоидным набором хромосом.
Отличительной особенностью первой стадии мейоза является то, что в период анафазы сами хромосомы не делятся на хроматиды, как при митозе, а лишь расходятся гомологичные пары хромосом к разным полюсам клетки и формируются две дочерние клетки с редуцированным наполовину набором хромосом, состоящим, однако, из двух хроматид.
Между первой и второй стадиями мейоза имеется непродолжительный период покоя — интеркинез, во время которого не Происходит репродукции хромосом.
Эквационное, или уравнительное, деление ^илогично митозу, где клетки последовательно проходят четыре Фазы: профазу II, метафазу II, анафазу II, телофазу П. На стадии °*шфазы II хромосомы разделяются на две хроматиды, которые затем с помощью нитей веретена расходятся к противоположным полюсам. На стадии телофазы II заканчивается формирование
21
 еще двух клеток. В результате после двух последовательных стадий мейоза из каждой клетки образуются четыре новые с гаплоидным набором хромосом. Для более наглядного представления всех этих событий можно воспользоваться схемой мейоза, приведенной на рисунке 7. еще двух клеток. В результате после двух последовательных стадий мейоза из каждой клетки образуются четыре новые с гаплоидным набором хромосом. Для более наглядного представления всех этих событий можно воспользоваться схемой мейоза, приведенной на рисунке 7.

Рис. 7, Схем» последомтелных стадий мейоза (по К. Свенсону и П. Уэбстеру, 1980):
А — лептонема, предшествующая конъюгации хромосом; Б — начало конъюгации на стадии зигонемы; В— пахинема; Г— диплонема; Д— метафаза I; E— анафаза I; Ж-телофаза I; 3 — интерфаза между двумя делениями мейоза; И— профаза II; К— метафаза II; Л— тело-фаза II
22
Таким образом, в результате двух меиотических делений из одной клетки с диплоидным набором хромосом образуются четыре клетки с гаплоидным набором хромосом и в 2 раза меньшим, чем в соматических клетках, содержанием ДНК. Вероятностный характер распределения материнских и отцовских гомологичных хромосом в разные клетки позволяет создать новые комбинации негомологичных хромосом в яйцеклетках и сперми-ях, чем достигается огромное число новых сочетаний наследственной информации.
Новые сочетания генетической информации возникают вследствие кроссинговера. Каждая из хромосом в метафазе I содержит участки, происходящие от отцовских и материнских хромосом. Рекомбинации хромосом при кроссинговере и вероятностное распределение их по клеткам — причины наследственной изменчивости организма. Мейоз, оплодотворение и митоз обеспечивают поддержание постоянства числа хромосом в смежных поколениях видов. В этом их биологическое значение.
Патология мейоза. Основная патология мейоза — нерасхождение хромосом- Оно может быть первичным, вторичным и третичным. Первичное нерасхождение возникает у особей с нормальным кариотипом. При этом на стадии анафазы I нарушается разделение бивалентов и обе хромосомы из пары гомологов переходят в одну клетку, что приводит к избытку хромосом в данной клетке (и + 1) и недостатку в другой (и—1). Вторичное нерасхождение возникает в гаметах у особей с избытком (трисо-мией) одной хромосомы в кариотипе. В результате этого в процессе мейоза образуются и биваленты, и униваленты. Третичные нерасхождения наблюдают у особей, имеющих структурные перестройки хромосом, например транслокации. Нерасхождение хромосом отрицательно влияет на жизнеспособность организма животных. Подробно об этом будет изложено в последующих главах.
Отличительная особенность мейоза у самок — образование в первом и втором меиотических делениях так назьгааемых полярных телец, которые впоследствии дегенерируют и в размножении не участвуют. Неравные деления в овогенезе обеспечивают яйцеклетке необходимое количество цитоплазмы и запасного желтка, чтобы она могла выжить после оплодотворения. В отличие от спермиогенеза, который у самцов происходит как во внутриутробный (пренатальный) период, так и после рождения (постна-тальный период), у самок яйцеклетка после рождения не образуется. Однако к концу пренатального периода у самок накапливается огромное количество овоцитов (у коров, например, десятки тысяч), а созревают и дают начало развитию потомства лишь немногие из них. Этот резерв генетического материала, особенно у малоплодных животных, таких, как крупный рогатый скот, в настоящее время начинают использовать для искусственного
23
стимулирования созревания многих яйцеклеток, последующего их оплодотворения и пересадки (трансплантации) специально подготовленным коровам-реципиентам. Таким образом от одной ценной коровы в год можно получить не одного, а несколько телят.
Оплодотворение наступает после слияния гаплоидных сперматозоида и яйцеклетки и образования диплоидной клетки — зиготы, дающей начало развитию эмбриона. При делении клеток эмбриона, содержащих хромосомы матери и отца, генетическая информация поступает во все клетки нового организма.
Морфологические и функциональные различия хромосомных наборов — основные причины эмбриональной смертности после оплодотворения при скрещивании разных видов или бесплодия гибридов. Так, при скрещивании зайца и кролика не происходит имплантации и развития оплодотворенной яйцеклетки. Эмбриональная смертность наступает при скрещивании козы и овцы. Мужские гибвиды осла (2 л = 62) и лошади (2л = 64), как известно, не дф.т потомства, они стерильные, или бесплодные. Стерильность гибридов-самцов наблюдается при скрещивании европейского крупного рогатого скота с бизоном, зубром, яком, а также гауром и гаялом.
Это связано с тем, что у гибридов нарушаются процессы спермиогенеза. Однако при скрещивании европейского крупного рогатого скота с азиатским горбатым зебу, дикого кабана (2л = 36) и домашней свиньи (2л = 38) потомство рождается плодовитое. В результате сложных вариантов скрещиваний в последнее время удалось получить плодовитых гибридов крупного рогатого скота с зуС^ом, бизоном, яком. Гибриды отличаются такими ценными качествами, как крепкое телосложение, высокая жизнеспособность, хорошие мясные признаки.
Контрольные вопросы. 1. Каковы морфологическое строение и химический состав хромосом? 2. Что такое кариотип и каковы его особенности у разных видов животных? 3. Что такое митоз и каково его биологическое значение? 4. Что такое гаметогенез и каковы его особенности у самцов и самок? 5. Что вы знаете о мейозе и его биологическом значении? 6. Каковы основные формы патологий митоза и мейоза?
|
|
|
 Скачать 5.3 Mb.
Скачать 5.3 Mb.