В. Н. Сайтаниди Рецензент членкорреспондент расхн в. Ф. Красота Петухов В. Л. и др. П31 Ветеринарная генетика В. Л. Петухов, А. И. Жигачев, Г. А. Назарова. 2е изд., перераб и доп. М. Колос, 1996. 384 с ил. Учебники
 Скачать 5.3 Mb. Скачать 5.3 Mb.
|

|
Глава 8 БИОТЕХНОЛОГИЯ Термин «биотехнология» получил широкое распространение в 70-е годы нашего столетия. Биотехнология опирается, с одной стороны, на древнейшие традиции бродильных и микробиологических производств, с другой — на новейшие открытия биологических наук. Биотехнология — это наука об использовании живых организмов и биологических процессов в производстве. Это комплексная многопрофильная область научно-технического прогресса, включающая многообразный микробиологический синтез, генетическую и клеточную инженерию, инженерную энзимоло-гию. Биотехнология возникла на стыке микробиологии, биохимии и биофизики, генетики и молекулярной генетики, цитологии и иммунологии. Уровень развития ее во многом определяет научно-технический потенциал страны. Стоимость мировой биотехнологической продукции к концу XX в., по прогнозам специалистов, достигнет 20 млрд долларов. Прогресс биотехнологии в животноводстве в предстоящие 10—15 лет будет определяться развитием генной, клеточной и эмбриогенетической инженерии. ГЕННАЯ ИНЖЕНЕРИЯ Генная инженерия — раздел биотехнологии, связанный с целенаправленным конструированием in vitro новых комбинаций генетического материала, способного размножаться в клетке и синтезировать определенный продукт. Генная инженерия решает следующие задачи:
Получение генов. Известны два способа искусственного синтеза генов вне организма — химический и ферментативный. Химическим путем в 1969 г. американский ученый Г. Корана с сотр. синтезировали ген аланиновой тРНК дрожжей. Этот ген включал 77 пар нуклеотидов, последовательность которых была 103 уже ранее расшифрована. Ученые сначала синтезировали фрагменты ДНК длиной от 5 до 12 нуклеотидов, затем соединили их в определенном порядке при помощи открытого к тому времени фермента лигазы. Однако ген аланиновой тРНК при введении в клетку кишечной палочки или бесклеточную среду не функционировал. Оказалось, что он не имел регуляторных элементов — промотора, где локализована точка инициации синтеза, и терминальных кодонов, которые дают сигнал о завершении синтеза иРНК. В 1976 г. Г. Корана с сотр. осуществили синтез гена супрессорной тирозиновой тРНК протяженностью 126 пар нуклеотидов. Были также синтезированы примыкающие к гену регу-ляторные участки: промотор (52 пары нуклеотидов) и терминатор (21 пара нуклеотидов) и прикрепленные к концам полимера тетрануклеотиды ААТТ и ТТАА. В этом случае искусственно синтезированный ген, встроенный в геном мутантного фага Т4, при введении в живую кишечную палочку оказался работоспособным^ В 1979 г. в нашей стране под руководством Ю. А. Овчинникова и М. Н. Колосова химическим путем с помощью ферментов были синтезированы гены гормонов человека и животных — энкефалина и брадикинина. Химико-ферментативный синтез довольно широко применяют в генной инженерии для получения мелких генов. Для получения генов животных, растений и человека, размер которых составляет 1000—3000 нуклеотидов, этот метод слишком сложен и пока неосуществим. Ученые нашли более простой способ получения таких генов. В 1970 г. Г. Темин с сотр. обнаружили фермент обратную транскриптазу (ревертазу). В 1972 г. было открыто, что некоторые онкогенные вирусы при помощи обратной транскриптазы могут синтезировать ДНК, используя в качестве матрицы иРНК. Дальнейшие исследования показали, что матрицами для образования копий ДНК могут служить не только РНК онкогенных вирусов, но и другие иРНК. Это открывало принципиальные возможности ферментативного синтеза любых индивидуальных генов (ДНК), используя их РНК-копии. Под ферментативным синтезом гена имеют в виду транскрибирование комплементарной нити ДНК (гена) на молекулах РНК в пробирке. Система для синтеза представляет собой раствор, в котором содержатся все четыре нуклеотида, входящих в состав ДНК, ионы магния, фермент обратная транскриптаза (ее получают из онкогенных вирусов) и матричная (информационная) РНК, кодированная геном, копию которого ставится задача снять. На иРНК обратная транскриптаза синтезирует комплементарную ей цепь ДНК, а затем на ней при помощи этого же фермента синтезируется вторая цепь ДНК. В результате получается ген по структуре такой же, как и тот, на котором была синтезирована иРНК. Этим способом в лабораториях многих стран создан целый 104 ряд генов. В нашей стране йод руководством В. А. Энгельгардта был разработан проект «Ревертаза» — программа синтеза генов с помощью этого фермента. В осуществлении проекта участвовали ведущие отечественные и зарубежные институты. В итоге с 1974 по 1978 г. были синтезированы гены глобина голубя, кролика и человека, а также гены митохондрий печени крыс, часть гена, кодирующего иммунные белки мышей, и др. Гены, синтезированные при помощи обратной транскриптазы, не имеют регуляторных участков и функционально неактивны. Поэтому транскрибирование копий ДНК рекомендуется проводить с про-мРНК, которые имеют все необходимые копии регуляторных частей гена. Кроме изложенных способов ген можно получить путем выделения с помощью трансдуцирующих фагов. Таким путем в 1969 г. был впервые выделен лактозный ген кишечной палочки. Однако такой способ получения генов не всегда пригоден, так как предусматривает строгие места локализации фагов. Поэтому используются и другие приемы выделения фрагментов ДНК с нужными для переноса генами. Рестриктирующие эндонуклеазы (рестриктазы). Важным событием для развития генной инженерии было открытие в клетках бактерий ферментов, способных разрезать молекулу ДНК в строго определенных местах. Ферменты эти называются рестрикти-рующими эндонуклеазами или рестриктазами, а процесс «разрезания» молекулы ДНК называется рестрикцией. С рестриктазами связаны дальнейшие успехи в молекулярной биологии. Они стали одним из главных элементов генной инженерии. Участок ДНК, узнаваемый определенной рестриктазой, включает специфическую последовательность из 6—8 пар оснований, являющихся палиндромом. Палиндромом называется последовательность ДНК, которая считывается одинаково в обоих направлениях, начиная от З'-конца каждой цепи. Например, рестриктаза E.coli под названием EcoRI узнает последовательность Г I ААТТ Ц Ц ТТАА t Г и, прикрепляясь к ней, делает по одному однонитевому надрезу с обеих сторон, т. е. разрезает ее в симметричных участках, указанных стрелками. В результате двухцепочная молекула ДНК, если была кольцевой, вследствие разрыва приобретает линейное строение. На краях молекулы образуются липкие концы, представленные однонитевыми участками из четырех нуклеотидов: на одном конце будет последовательность ААТТ, на другом — ТТАА. При наличии липких концов молекула ДНК из линейной формы вновь способна замкнуться в кольцо без дополнительной обработки. Были обнаружены рестриктазы, узнающие самые разнообразные последовательности нуклеотидов. Например, рестриктаза EcoRII узнает последовательность ЦЦТГГ. 105 К настоящему времени известно свыше 200 рестриктаз, характерных для разных видов микроорганизмов. Это открывает новые возможности для экспериментаторов. Огромные молекулы высших организмов включают большое число мест разрезания для ферментов рестрикции. При обработке ДНК рестриктазами образуются многочисленные фрагменты ДНК, в которых представлены отдельные гены. Затем гены можно соединять в определенные структуры. Остающиеся в такой структуре разрывы нитей ДНК воссоединяются лигазой. Имеется и другой способ получения фрагментов ДНК с липкими концами. Он состоит в том, что выделенные или искусственно синтезированные участки ДНК обрабатывают эндонуклеазой, укорачивающей участки ДНК с обоих концов, после чего при помощи фермента полинуклеотидтран-сферазы пристраивают к этим концам последовательности адени-ловых и тимидиловых нуклеотидов. Длина липких поли-А и поли-Т составляет 50—100 нуклеотидов. При встраивании гена в вектор используются оба рассмотренных метода и часто совместно. Рекйибинантные ДНК. Рекомбинантная ДНК — это искусственно полученная молекула ДНК. Она имеет форму кольца, включает ген (гены), составляющий объект генетических манипуляций, и так называемый вектор, обеспечивающий размножение рекомбинантной ДНК и синтез в клетке хозяина определенного продукта, кодируемого внесенным геном. Векторами являются те компоненты рекомбинантных ДНК, которые способны акцептировать чужеродные гены и обеспечивать их репликацию в клетках хозяина. Векторы должны обладать следующими особенностями: 1) иметь свойства регогакона; 2) нести субстратные участки для рестриктаз, иначе невозможна встройка ДНК; 3) содержать один или несколько маркирующих генов, чтобы по фенотипу можно было определить факт его передачи. Исследования показали, что эффективными векторами являются плазмиды. Из них в качестве векторов используются Col El, pSC 101 и др. Из вирусов в качестве векторов используют фаги X, SV 40 и их производные. Векторы придают рекомбинантной молекуле способность воспроизводиться независимо от хромосомы клетки бактерии. Соединение вектора с фрагментом ДНК может производиться следующими путями: при помощи липких концов, образующихся в ДНК под действием эндонуклеаз рестрикции; дополнительного синтеза полинуклеотидных фрагментов каждой из цепей ДНК (поли-А и поли-Т); соединения тупых концов при помощи Т4-лигазы. На рисунке 25 показаны ферментативный синтез гена и встраивание его в векторную плазмиду. Слева на иРНК при помощи обратной транскриптазы синтезируется цепь ДНК (кДНК), затем иРНК удаляют щелочью и при помощи ДНК-полимеразы достраивают вторую цепь кДНК, экзонуклеазой укорачивают обе цепи ДНК и концевой трансферазой пришивают к их концам поли-Т-последовательности. На этом же рисунке 106 ПЛАЗМИДА мРНК ОБРАТНАЯ ТРАНСКРИПТАЗА РЕСТРИКТАЗА , ЛИНЕЙНАЯ \ ДНК ' ПЛАЗМИДЫ мРНК «ДНК ЭКЗОНУКЛЕАЗА ДНК - ПОЛИМЕРАЗА ДВУХНИТЕВАЯ кДНК ЭКЗОНУКЛЕАЗА КОНЦЕВАЯ ТРАНСФЕРАЗА АААА- •АААА КОНЦЕВАЯ ТРАНСФЕРАЗА ГИБРИДНАЯ МОЛЕКУЛА СО ВСТРОЕННЫМ ГЕНОМ РАЗРЫВЫ УСТРАНЯЮТСЯ ЛИГАЗОЙ Рис. 25. Ферментативный синтез гена и встраивание его в векторную плазмиду (по С. М. Гершензону) справа показано, что кольцевую ДНК векторной плазмиды разрезают рестриктазой и превращают ее в линейную форму, затем экзонуклеазой цепи укорачивают и концевой трансферазой пришивают к ним поли-А-последовательности. На последнем этапе соединяют оба типа молекул ДНК и получают гибридную молекулу — векторную плазмиду со встроенным в нее синтезированным геном. Разрывы в цепях ДНК воссоединяют лигазой. В данном случае было известно, какой ген включен в векторную плазмиду. Но в работах по трансгенозу чаще имеют дело со многими фрагментами ДНК, и среди них только единичные включают ген, который подлежит переносу. Для получения фрагментов ДНК с определенным геном применяют так называемый способ дробового ружья, который заключается в том, что ДНК механически или ферментами дробится на множество мелких фрагментов, после чего их вслепую гибридизируют с молекулами ДНК вектора. Перед этим ДНК вектора обрабатывают рестриктазой для придания им линейной формы и образования липких концов. После введения рекомбинантных молекул в кишечную палочку при помощи селективных сред выделяют те бактерии, в которые попал фрагмент ДНК с нужным геном. Включенный ген обнаруживают по продукту его действия в виде определенного вещества (фермент, гормон и т. д.). Размножение в бактериях 107 Генетический код ДНК лактозного оперона I Lac .Р .0. р-гал I Химический синтез ДНК Ген соматостатина идентичных рекомбинантных ДНК называется клонированием. Каждый клон бактерий содержит свою рекомбинантную ДНК. Разрабатываются методы, позволяющие производить замены нуклеотидов в ДНК клонированного гена и тем самым изменять свойства кодируемого этим геном белка. ?■ ФРАГМЕНТЫ ДНК, xj ОБРАБОТАННЫЕ РЕСТРИКТАЗОЙ Л 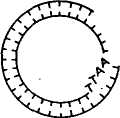 В 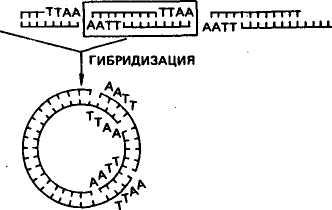 ведение в клетку рекомбинантных молекул и синтез чужеродного белка. Чаще всего рекомбинантные молекулы вводятся в клетки бактерий методом трансформации. Сначала клетки бактерий с целью повышения их способности поглощать плазмидную ДНК обрабатывают хлористым кальцием или хлористым барием. После этого в клеточную взвесь вводят раствор с рекомбинантны-ми молекулами. Некоторые из этих молекул проникают внутрь клетки, и часть из них, прикрепясь к мембранам клетки, начинает размножаться и функционировать (рис. 26). ведение в клетку рекомбинантных молекул и синтез чужеродного белка. Чаще всего рекомбинантные молекулы вводятся в клетки бактерий методом трансформации. Сначала клетки бактерий с целью повышения их способности поглощать плазмидную ДНК обрабатывают хлористым кальцием или хлористым барием. После этого в клеточную взвесь вводят раствор с рекомбинантны-ми молекулами. Некоторые из этих молекул проникают внутрь клетки, и часть из них, прикрепясь к мембранам клетки, начинает размножаться и функционировать (рис. 26).ПЛАЗМИДА_pSC_101.__1_ДЕЙСТВИЕ_ДНК-ЛИГАЗЫ'>ПЛАЗМИДА pSC 101. 1 ДЕЙСТВИЕ ДНК-ЛИГАЗЫ 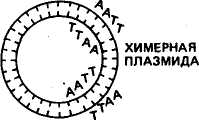 ТРАНСФОРМАЦИЯ ХРОМОСОМА ТРАНСФОРМИРОВАННАЯ БАКТЕРИЯ 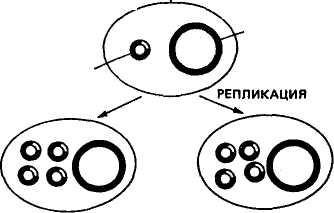 ПЛАЗМИДА ОБРАБОТАННАЯ РЕСТРИКТАЗОЙ ТРАНСФОРМИРОВАННЫЕ ДОЧЕРНИЕ КЛЕТКИ Рис. 26. Типичный опыт генетической инженерии (по Beckingeham—Smith, 1975) 108 4аАГщ|аТГ| ГЦТ ГГТ ТГТ ААГ ААЦ ТТЦ ТТТ Т Г I I I {ЦТАГ | ГАТ АГТ | ТГТ ГЦТ ТЦА ШТ Щ Г А tin vivo Som -Ала-Гли-Цис-Лиз -Асн -Фен -Фен—^ S Д  НК плазмиды pBR 322 НК плазмиды pBR 322Р-гал NH2- Трп Лиз НО-Цис-Сер-Тре -Фен -Тре-обработка цианогенбромидом фрагменты Д-г<м+М2-Ала-Гли-Цис-Лиз-Асн-Фен-Фен->. S Трп S Лиз НО-Цис-Сер-Тре-Фен-Тре-^ Активный соматостатин Рис. 27. Функционирование химически синтезированного гена соматостатина человека в клетках Е. coli (по К. Итакуре и др.) Одно из наиболее важных для практики направлений генной инженерии — конструирование микроорганизмов-продуцентов нужных веществ. Первыми в этом направлении были исследования К. Итакуры и Г. Боейра с соавт. (1977). Им удалось добиться экспрессии гена, кодирующего гормон соматостатин в клетках кишечной палочки (рис. 27). Затем в разных странах клетки кишечной палочки использовались для синтеза ряда белков и гормонов человека и животных: инсулина, интерферонов, гормона роста, урокиназы, кальцитонина, альбумина, тимозина и др. В лабораториях А. А. Баева, Ю. А. Овчинникова, М. Н. Колосова, Е. Д. Свердлова и др. осуществлена экспрессия генов, кодирующих энкефалин, лейкоцитарный интерферон, брадикинин, сома-тотропин и др. В последние годы уделяется много внимания созданию генно-инженерных вакцин. Получают антигены из рекомбинантных микроорганизмов или культур клеток, в которые введен определенный ген возбудителя болезни. Этим методом получен материал для вакцинации против гепатита В, гриппа А, малярии, ящура, бешенства, паравируса свиней и др. В нашей стране произведена обратная транскрипция РНК вируса гепатита А. ДНК, кодирующая вирусный белок гепатита, была пришита к 109 ДНК вируса оспы. В результате ослабленная культура оспы вызывает иммунитет против опасной болезни — гепатита. Вакцина проходит производственное испытание. Штаммы бактерий, продуцирующие вещества, активные в организме человека и животных, могут быть использованы для промышленного производства лекарственных препаратов. В нашей стране ведутся работы по получению методами генной инженерии суперпродуцентов продуктов, свойственных клеткам, таких, как аминокислоты, витамины, ферменты, а также по созданию культур, активно разлагающих нефть, пластмассы, разрушающих нафталин, фиксирующих азот, и т. д. КЛЕТОЧНАЯ ИНЖЕНЕРИЯ Под клеточной инженерией понимают метод конструирования клеток нового типа на основе их культивирования, гибридизации и реконструкции. К у*л ь т у р а клеток — метод сохранения жизнеспособности клеток вне организма в искусственно созданных условиях жидкой или плотной питательных сред. Использование культуры клеток начато в 50-е годы нашего столетия, когда была показана возможность выращивания вирусов в культивируемых клетках. Затем развитие вирусологии и методов культивирования клеток животных и человека позволило ученым приступить к созданию вирусных вакцин. Для культивирования могут быть использованы клетки опухолевых тканей, клетки различных органов, лимфоциты, фибро-бласты, эмбрионы, клетки почек животных и человека, раковые клетки человека и т. д. Культуры, приготовленные непосредственно из тканей организма, называются первичными. В большинстве случаев клетки первичной культуры можно перенести из культуральной чашки и использовать для получения вторичных культур, которые можно последовательно перевивать в течение недель и месяцев. Многие клетки при этом сохраняют признаки дифференцировки тех тканей, из которых они были получены. Например, фибробласты продолжают секретировать коллаген, клетки скелетных мышц эмбриона сливаются и образуют гигантские мышечные волокна, которые спонтанно сокращаются в чашке для культуры тканей, и т. д. Так как все это происходит в культуре, то является доступным для изучения с помощью приемов, которые неприемлемы при работе с интактными тканями. Клетки животных и человека выращивают на специальных средах в виде суспензии или монослоя на стекле. Технология культивирования некоторых клеток животных настолько хорошо отработана, что может быть использована в производственных целях для получения различных продуктов. В настоящее время клонировано много генов, кодирующих синтез белков разной 110 биологической ценности. Некоторые из таких генов удалось перенести в клетки животных, и они стали продуцентами биологически активных белков. В промышленных масштабах в биореакторах с использованием клеток животных налажено производство таких белков. Они используются как медицинские препараты. Например, эритропоэтин (гормон, стимулирующий образование красных кровяных тел), активатор плазминогена (используется для предотвращения образования тромбов), фактор свертывания крови III (используется при гемофилии), инсулин (для лечения диабета), поверхностный белок вируса гепатита В, интерлейкины и др. |