Генетика. Вопросы к экзамену. Генетика. Вопросы. Вопросы для сдачи экзаменов по дисциплине Общая и медицинская генетика
 Скачать 1.29 Mb. Скачать 1.29 Mb.
|

|
Диспергированные повторяющиеся последовательности чаще всего разбросаны по геному по отдельности. Согласно размеру, их подразделяют на две группы: длинные диспергированные элементы, LINE, и короткие, SINE. Оба класса являются ретротранспозонами. Умеренно повторяющиеся последовательности – представлены в геноме десятками или несколькими сотнями копий и участвуют в осуществлении жизненно важных функций. Это гены гистонов, тРНК, рРНК; их еще называют генами «домашнего хозяйства». Большую группу умеренно повторяющихся последовательностей составляют подвижные генетические элементы. Они могут быть расставлены по всему геному или собраны в группы. Высокоповторяющиеся последовательности – свыше 1000 повторов, представлены сателлитными (спутниковыми) ДНК. Они состоят из коротких тандемных повторов, организованных в блоки. Мини- и микросателлитные тандемные повторы разбросаны по всему геному и представляют собой уникальную для каждой особи комбинацию. У человека эти повторы используют в судебно-медицинской практике. Регуляция активности генов. Уровни регуляции. Система оперона по Жакобу и Моно. Регуляторные белки. Перестройки генома. Подвижные элементы генома. Не все белки в данный момент нужны организму – происходит регуляция генов. У эукариот нет оперонов, нет единой регулирующей системы, есть регуляция ферментами, один регулятор для многих генов, у генов много регуляторов, так как компоненты одной биохимической реакции разбросаны по разным хромосомам, есть энхансеры и сайленсеры. Кроме того, необходимо формирование особых сигнальных путей, передающих сигнал с поверхности клетки или из цитоплазмы в ядро. Индукция или репрессия осуществляется с помощью факторов внешней среды (света, тепла) и внутренней (гормонов, транскрипционных факторов). Можно рассказать про стероиды и другие гормоны: как действуют. Гормон-индуцируемая экспрессия опосредуется специфическими гормональными респонсивными элементами (hormone response elements (HRE)). Они расположены вблизи генов, которые ими регулируются, и служат для связывания специфических белков, которые действуют как транскрипционные факторы. Также экспрессия может регулироваться белками, которые распознают специфические последовательности ДНК в результате точного соответствия их поверхности двойной спирали ДНК. В их число входят белки типа «цинковый палец», «лейциновая застёжка», «спираль – петля – спираль». Они или взаимодействуют с большой бороздкой, изменяя конформацию ДНК и открывая доступ РНК-полимеразам, или с другими белками, что тоже способствует транскрипции. Схематично генная экспрессия может регулироваться на четырех различных уровнях: уровень инициации транскрипции: образование гетерохроматина – конститутивного (повторяющиеся последовательности, мигрирующие элементы) и факультативного, ремоделирование в случае надобности (модификация и замещение гистонов: метилирование, ацетилирование, фосфорилирование, убиквитинирование); метилирование ДНК, формирование молчащей Х-хромосомы; геномный импринтинг (экспрессируются только те гены, которые наследовались от определённого родителя). уровень транскрипции: регуляция гормонами, РНК-интерференция (дцРНК нехарактерны для нормальных клеток, но они являются обязательным этапом жизненного цикла многих вирусов. Специальный белок Dicer, обнаружив в клетке дцРНК, «режет» её на небольшие фрагменты. Антисмысловая цепь такого фрагмента, которую уже можно называть малой интерферирующей РНК, связывается с комплексом белков под названием RISC, центральный элемент которого – эндонуклеаза семейства Argonaute. Связывание с миРНК активирует RISC и запускает в клетке поиск молекул ДНК и РНК, комплементарных «шаблонной» миРНК. Судьба таких молекул – быть уничтоженными или инактивированными комплексом RISC), энхансеры и сайленсеры, редактирование ДНК (превращение Ц в У и образование стоп-кодона). уровень трансляции (РНК-и, сродство к факторам инициации и рибосомам) и уровень активности белка (аллостерическая регуляция, ингибирование, активирование). Механизм регуляции экспрессии генов у прокариот рассматривается на уровне оперона. Оперон – это группа регулируемых структурных генов, кодирующих ферменты. В состав оперона входят несколько структурных генов, гены, кодирующие рРНК и тРНК, а также общая регуляторная область – промотор, оператор и терминатор транскрипции. Ген-регулятор имеет собственный промотор и может находиться на некотором расстоянии от структурных генов. Он обеспечивает синтез белка-репрессора, который, связываясь с оператором, тормозит экспрессию. Оператор – участок ДНК, примыкающий к структурным генам оперона и управляющий их функцией. Если оператор свободен, то транскрипция разрешена, если он связывается с белком-репрессором, то работа этих генов прекращается. Промотор – участок ДНК, который либо непосредственно примыкает к оператору, либо перекрывается с ним. Есть позитивная и негативная регуляция: позитивная индукция: цАМФ соединяется с белком-активатором катаболического гена CAP, тот меняет конформацию и соединяется с ДНК, облегчая присоединение РНК-полимеразы; негативная индукция: появление лактозы приводит к тому, что репрессор связывается с ней, а не с геном, поэтому запускается транскрипция; негативная репрессия: появление триптофана или гистидина заставляет их связываться с репрессором, не имеющим сродства к оператору, менять конформацию и тем самым тормозить транскрипцию. Перестройки запланированные: У прокариот есть гены, отвечающие за размножение или образование спор, они разорваны, в необходимый момент ДНК вырезается, гены сшиваются и начинают работать. У эукариот в течение дифференцировки клеток могут вырезаться кластеры генов, например, в процессе пролиферации и дифференцировки В-лимфоцитов. У эритроцитов, тромбоцитов и клеток хрусталика глаза и вовсе удаляется ядро. Так уничтожается ненужная ДНК из дифференцированных клеток. Незапланированные – с помощью мобильных элементов. Количество мобильных элементов до 1000 (до 10% генома у человека), длина от 1 тыс. до 10 тыс. п. н., наличие на концах длинных концевых повторов – типичная черта строения мобильных элементов. Могут вырезаться и вставляться, могут транскрибироваться и встраиваться. Многие точечные мутации у дрозофилы, мыши и других организмов в действительности представляют собой результат встраивания или вырезания мобильных генетических элементов. Экспрессия генов. Этапы синтеза белка: транскрипция, процессинг, трансляция. Клинические примеры нарушений. Понятие «один ген-один фермент» и его условность. Биохимическая индивидуальность организма. Экспрессия генов – процесс передачи наследственной информации от гена к функциональному продукту. Этапы: транскрипция, сплайсинг РНК и стадия посттрансляционных модификаций, трансляция. Регуляция экспрессии позволяет контролировать собственную структуру и функцию и является основой дифференцировки клеток, морфогенеза и адаптации. Транскрипция начинается в определённых сайтах ДНК – промоторах и завершается в сайтах терминации. Гены эукариот состоят из двух структурных областей: кодирующей и регуляторной, которая запускается и контролирует работу РНК-полимеразы II, а через нее – синтез мРНК. Регуляторная область у эукариот включает промоторы, связывающие РНК-полимеразу, терминаторы, энхансеры (усилители транскрипции) и сайленсеры (ослабители). Энхансеры и сайленсеры могут входить в структуру генов, но чаще всего располагаются на расстоянии, служат местами для узнавания и связывания с регуляторными белками. У эукариот процесс транскрипции и последующее созревание мРНК протекает в клеточном ядре, он значительно усложнен. Во-первых, у эукариот функционируют три разные полимеразы, во-вторых, РНК-полимераза эукариот не может сама инициировать транскрипцию. Для ее активации необходимо большое количество белков, они называются факторами транскрипции. Общие факторы транскрипции объединяются в комплексы. В-третьих, регуляторные элементы у эукариот могут влиять на скорость транскрипции, даже если они расположены за тысячи пар нуклеотидов от промотора. Например, энхансеры образуют петлю, взаимодействуя с промотором, который непосредственно взаимодействует с факторами транскрипции и с РНК-полимеразой. В-четвёртых, транскрипция и трансляция разделены во времени и пространстве. Транскриптон – участок ДНК между промотором и сайтом терминации. Факторы транскрипции – белки, взаимодействующие с определёнными регуляторными сайтами ДНК, инициирующие, ускоряющие, замедляющие или блокирующие транскрипцию конкретного гена. РНК-полимераза I синтезирует пре-рРНК, РНК-полимераза II – пре-мРНК, РНК-полимераза III – пре-тРНК. 3 стадии: инициация, элонгация и терминация. ТАТА-фактор, ТАТА-бокс, раскручивание примерно одного витка спирали, синтез олигонуклеотида из 8-10 нуклеотидов, факторы инициации. Факторы элонгации, расхождение цепей, продвижение РНК-полимеразы, восстановление спирали ДНК. Сайт терминации, фактор терминации, отделение первичного транскрипта и РНК-полимеразы. Ковалентные модификации (модификации концов и сплайсинг). Процессинг – совокупность биохимических реакций, приводящих к преобразованию первичного транскрипта в зрелую мРНК. Процессинг у прокариот имеет весьма ограниченный характер и в основном затрагивает предшественников рРНК и тРНК, тогда как мРНК используется для трансляции сразу после их синтеза, еще до окончания их транскрипции. Посттранскрипционные модификациии РНК у эукариот связаны с копированием, полиаденированием и с экзон-интронным строением генов, а также прерывистым строением их гена, содержащего экзоны и интроны, что требует четкой системы сплайсинга первичных транскриптов. Информационная РНК может содержать от одного до десятков интронов, размеры которых сильно варьируются. В число компонентов, катализирующих процесс сплайсинга, входят мяРНК и десятки белков, обладающих ферментативной активностью. Они образуют мяРНП. Место соединения интрон/экзон узнается мяРНП, которые образуют более крупный комплекс, называемый сплайсосомой. Механизм сплайсинга включает расщепление и прикрепление 5'-конца интрона к 3'-концу аденина сплайсосомы, при этом образуется петлевая структура, называемая «лассо». 3'-конец экзона затем обычно замещает 3'-конец интрона, два экзона соединяются вместе. Альтернативный сплайсинг позволяет организму синтезировать разные по структуре и свойствам белки на базе одного гена. В формировании альтернативных мРНК задействованы разные механизмы. Первый из них состоит в том, что для образования различных мРНК могут использоваться разные промоторы. В этом случае образуются транскрипты, имеющие разное количество экзонов. Второй тип имеет место при изменении сайта полиаденилирования первичного транскрипта. Третий тип включает вырезку различных экзонов из одинаковых иРНК. При этом для формирования зрелых РНК могут использоваться различные экзоны. Трансляция: Большая СЕ (60S: 5S, 28S и 5,8S и до 50 белков), малая СЕ (40S: 18S РНК и более 30 белков). Три участка связывания – один для мРНК и два для тРНК. Малая СЕ присоединяет мРНК и декодирует информацию с помощью тРНК, большая СЕ ответственна за образование пептидных связей. Связывание мРНК у прокариот происходит в районе желобка. Главную роль здесь играет комплементарное взаимодействие пурин-богатого участка мРНК, так называемая последовательность Шайна-Дальгарно, с локализованной в 30S субчастице короткими, богатыми пиримидином участками З'-конца молекулы 16S рРНК. У эукариот присоедниняется кэп и АУГ-кодон. Экспрессия генов. Этапы синтеза белка: транскрипция, процессинг, трансляция. Понятие «один ген – один фермент» и его условность. См. выше Мутации. Классификации мутаций. Роль мутаций в эволюции. Мутации могут происходить под влиянием как внешних, так и внутренних воздействий. Генеративные и соматические; точечные, или генные, хромосомные, геномные мутации. 1901 г., де Фриз. Свойства: возникают внезапно, и мутировать может любая часть генотипа; чаще бывают рецессивными и реже – доминантными; могут быть вредными (большинство), нейтральными и полезными (очень редко) для организма; передаются из поколения в поколение; представляют собой стойкие изменения наследственного материала; м 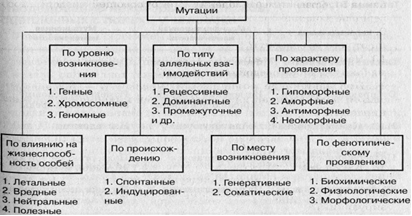 огут повторяться. огут повторяться.Точковые мутации – замена, выпадение или вставка одного или нескольких нуклеотидов. По эффекту бывают нонсенс-, сайленс- и миссенс-мутации, со сдвигом или без сдвига рамки считывания. Мутации могут вести к нарушению сплайсинга (сшивания экзонов и вырезания интронов). Отдельный тип мутаций — «динамические» мутации, которые характеризуются нарастанием триплетных повторов в кодирующих и некодирующих участках гена. Самой известной заменой (субституцией) является замена аденина (А) на тимин (Т) в 6-м кодоне гена α-глобина (ГАГ заменяется на ПТ), что приводит к замене аминокислоты глутамина на валин в полипептидной цепи α-глобина. Гомозиготные носители таких аллелей известны как лица, страдающие серповидноклеточной анемией. Наряду с серповидно-клеточной анемией существуют и другие мутации гена, приводящие к появлению аллелей, гомозиготные носители которых страдают β-талассемией. Хромосомные мутации – возникают из-за нарушения целостности хромосом. Разрывы хромосом происходят в ходе кроссинговера. Это ведет к появлению новых групп сцепления из-за делеции, дупликации или транслокации. При этом изменяется число генов в группе сцепления. Нарушение целостности хромосомы может сопровождаться инверсией из-за двух разрывов. В зависимости от того, включает ли данный участок область центромеры или нет, различают перицентрические и парацентрические инверсии. Фрагмент хромосомы без центромеры, отделившийся при разрыве, может быть утрачен клеткой при очередном митозе. Также он может прикрепляться к какой-то негомологичной хромосоме. Нередко две поврежденные негомологичные хромосомы взаимно обмениваются оторвавшимися участками – реципрокная транслокация. Присоединение фрагмента к своей хромосоме в другом месте – транспозиция. Иногда негомологические хромосом объединяются в одну хромосому – робертсоновская транслокация. Геном клетки – это половинное количество ДНК, включая ядерную и цитоплазматическую ДНК. К геномным мутациям относят изменение числа геномов или числа отдельных хромосом: анеу-, поли- и гаплоидии. Гаплоидия – это уменьшение числа хромосом вдвое, т. е. соматические клетки содержат одинарный набор хромосом (n). Гаплоидные организмы мельче, у них проявляются рецессивные гены, они бесплодны. Полиплоидия – увеличение числа хромосом, кратное гаплоидному набору в клетке. Встречаются триплоидные (Зn), тетраплоидные (4n), пентаплоидные (5n) организмы. Полиплоидия может возникнуть в результате нарушения расхождения хромосом при митозе, слияния клеток соматических тканей либо их ядер, нарушений мейоза, приводящих к образованию гамет с нередуцированным числом хромосом. У человека более 20% всех спонтанных абортов – это триплоидный набор хромосом в кариотипе. Такие особи погибают в первые часы жизни или дни после рождения. Тетраплоидию у человека наблюдают только в материале спонтанных абортов. Анеуплоидия – изменение числа хромосом в клетках организма за счет потери (моносомия) или добавления (полисомия) отдельных хромосом. Механизм анеуплоидии связан с нарушением расхождения хромосом при мейозе. Процесс образования мутаций во времени и пространстве называется мутагенезом. Вещества, вызывающие мутации в клетках – мутагены. В зависимости от происхождения различают спонтанные и индуцированные мутации. Спонтанные мутации возникают в нормальных природных условиях без видимых причин как ошибки при воспроизведении генетического материала, поскольку редупликация не происходит с абсолютной точностью. Они возникают в условиях природного радиоактивного фона Земли в виде космического излучения, радиоактивных элементов, радионуклидов в клетках организмов. Спонтанная мутация может возникнуть в любой период индивидуального развития и поразить любую хромосому или ген. Частота встречаемости 1:100000. Индуцированные – возникают в результате действия мутагенов. Поэтому по типу клеток различают генеративные и соматические мутации. Большая часть канцерогенных веществ вызывает мутации в соматических клетках. Возникшие повреждения в ДНК могут бесследно исчезнуть благодаря репарации. Проявление мутантного гена может быть подавлено действием другого гена. Группы сцепления. Кроссинговер и его биологическая роль. Основные положения хромосомной теории наследственности: Гены локализованы в хромосомах. Хромосомы содержат неодинаковое число генов, набор генов каждой из негомологичных хромосом уникален. Аллельные гены занимают одинаковые локусы в гомологичных хромосомах. Гены расположены в хромосоме в линейной последовательности. Гены одной хромосомы образуют группу сцепления. Число групп сцепления – n (n+1, если гетерогаметный пол, как у мужчин), поскольку группу сцепления составляют две гомологичные хромосомы, в которых локализованы одинаковые гены. Сцепление нарушается в результате кроссинговера, частота которого прямо пропорциональна расстоянию между генами в хромосоме. Каждый биологический вид характеризуется определенным набором хромосом – кариотипом. Независимое комбинирование признаков (третий закон Менделя) осуществляется при условии, что гены, определяющие эти признаки, находятся в разных парах гомологичных хромосом. Совместное наследование генов Т. Морган предложил назвать сцепленным наследованием. Он определил, что гены находятся в хромосомах, что все гены из одной хромосомы образуют группу сцепления и наследуются совместно, что в отдельных случаях из-за явления кроссинговера может наблюдаться неполное сцепление генов. Например, в опытах с дрозофилой полное сцепление наблюдается только у самцов. Как показали опыты, у самки дрозофилы и других представителей мужского и женского пола полного сцепления нет, а наблюдается частичное сцепление. В потомстве от дигетерозиготной самки дрозофилы и рецессивной гомозиготы было получено 4 типа особей, но соотношение было не 1 :1 : 1 : 1, а 41,5 : 8,5 : 8,5 : 41,5. Опираясь на наблюдения Ф. Янсена, обнаружившего в мейозе хиазмы, Морган предположил, что особи Bbvv и bbVv появились в результате перекреста и разрыва гомологичных хромосом, благодаря чему и происходит обмен гомологичными участками гомологичных хромосом в точках между генами В и V. |