Генетика. Вопросы к экзамену. Генетика. Вопросы. Вопросы для сдачи экзаменов по дисциплине Общая и медицинская генетика
 Скачать 1.29 Mb. Скачать 1.29 Mb.
|

|
Способы деления клеток: Амитоз – прямое, простое деление клетки (неполноценное). Митоз – сложное, непрямое, полноценное деление клетки. Мейоз – сложное, непрямое, редукционное деление клеток репродуктивных органов. Ядро клетки и хромосомы. Свойство хромосом и понятие о кариотипе. Первые подробные описания морфологии хромосом и их перемещений во время деления клеток были описаны российским ученым Чистяковым при изучении спор плаунов. Число и форма хромосом являются видовым признаком. Морфологически хромосомы различаются между собой размерами и формой, которая определяется величиной их плеч относительно центромеры. Классификацию хромосом разработал русский ученый Навашин в 1912-1914 гг. Варианты строения: мета-, субмета-, акро- и телоцентрические, со вторичной перетяжкой и спутниками. Хромосомы состоят из ДНК и белков. На долю белков приходится около 60%, на ДНК – 40% общей массы. Белки подразделяются на гистоновые и негистоновые. Гистоновые представлены пятью основными фракциями, составляют от 40 до 80% всех белков, состоят из небольших молекул, имеющих положительные электрические заряды, поскольку в их составе преобладают аргинин и лизин. Полагают, что гистоновые белки выполняют важную функцию, связанную со специфической упаковкой огромной по размерам молекулы ДНК в компактную структуру хромосомы. Негистоновые белки выполняют разные функции: участвуют в процессах репликации ДНК, транскрипции, репарации. Хромосомы состоят из центромер и теломер. Теломеры образуются из множественных 68 нуклеотидных повторов из блоков ТТАГГГ и играют важную роль в создании специфической архитектуры и внутренней упорядочности клеточного ядра. Они предотвращают слипание концов хромосом и прикрепляются к специальной внутриядерной структуре. При каждом раунде репликации хромосомы будут укорачиваться на 1020 нуклеотидов (все зависит от размера РНК-затравок) и в первую очередь сокращается длина теломер. В случае репликации кольцевых бактериальных ДНК этой проблемы не существует. Проблема недорепликации З'-концов линейных молекул решается эукариотической клеткой с помощью специального фермента – теломеразы. Этот фермент был обнаружен в 1985 г. у инфузорий, а затем в дрожжах, растениях и животных. Теломераза – это ДНК-полимераза, достраивающая те самые 68 нуклеотидов. Помимо белковой части теломераза содержит РНК, играющую роль матрицы для наращивания ДНК-цепи. У хромосом большинства типов соматических клеток теломеры укорачиваются с возрастом, и это считают одним из важных факторов, определяющих продолжительность жизни особи. Таким образом, изучение механизма работы теломеразы и регуляции ее экспрессии в клетке помогут понять молекулярные основы процесса старения и злокачественной трансформации клеток. Еще одно доказательство связи длины теломеры со старением у человека было получено при изучении людей с прогерией. В соответствии с гипотезой уменьшения длины теломер в процессе старения соматические клетки людей с прогерией имеют короткие теломеры и проявляют сниженную способность к росту и размножению. В области первичных перетяжек располагается специфическая центромерная ДНК с часто повторяющимися короткими последовательностями нуклеотидов ГЦ, АТ, с которыми взаимодействуют специальные белки кинетохоров. В 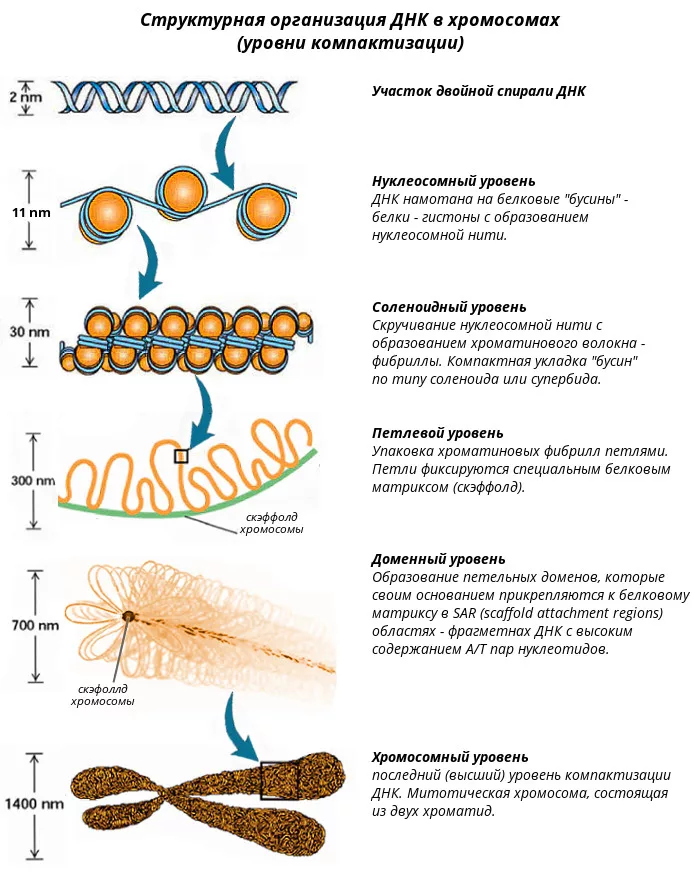 околоцентромерных участках и коротких плечах акроцентрических хромосом прокрашивается гетерохроматин, обозначаемый как конститутивный, который постоянно выявляется и во время митотического деления клетки и в интерфазном ядре. Полагают, что в ДНК-структурированном районе гетерохроматина гены, отвечающие за синтез белка, либо отсутствуют, либо таких генов мало. Другой тип гетерохроматина — факультативный, возникает путем компактизации эухроматиновых районов и содержит гены, участвующие в метаболизме белка. Конденсация факультативного гетерохроматинового района хромосом носит обратимый характер, выражающийся в возможности деконденсации этих районов. У млекопитающих и человека в соматических клетках женского организма одна из двух Х-хромосом находится в состоянии компактизации (тельце Барра), это и есть пример факультативного гетерохроматина. околоцентромерных участках и коротких плечах акроцентрических хромосом прокрашивается гетерохроматин, обозначаемый как конститутивный, который постоянно выявляется и во время митотического деления клетки и в интерфазном ядре. Полагают, что в ДНК-структурированном районе гетерохроматина гены, отвечающие за синтез белка, либо отсутствуют, либо таких генов мало. Другой тип гетерохроматина — факультативный, возникает путем компактизации эухроматиновых районов и содержит гены, участвующие в метаболизме белка. Конденсация факультативного гетерохроматинового района хромосом носит обратимый характер, выражающийся в возможности деконденсации этих районов. У млекопитающих и человека в соматических клетках женского организма одна из двух Х-хромосом находится в состоянии компактизации (тельце Барра), это и есть пример факультативного гетерохроматина.Свойства хромосом: индивидуальное строение; парность в соматических клетках; постоянство числа; способность к самовоспроизводству. Первый уровень компактизации ДНК – нуклеосомный. Играет регуляторную и структурную роль, обеспечивая плотность упаковки ДНК в 6-7 раз. Второй уровень – фибриллярный. В митотических хромосомах и в интерфазных ядрах выявляются фибриллы хроматина с диаметром 25-30 нм. В результате такой упаковки возникает фибрилла спирального типа с центральной полостью. Нуклеомерный уровень укладки хроматина обеспечивает 40-кратное уплотнение ДНК. Нуклесомный и нуклеомерный (супербидный) уровни компактизации ДНК хроматина осуществляются за счет гистоновых белков. Третий уровень – хромомерный. В высших уровнях организации хроматина специфические белки связываются с особыми участками ДНК, которая в местах связывания образует большие петли или домены. Четвёртый – хромонемный. Образуется из петель, собранных в розетку. Пятый – хромосомный. Набор индивидуальных хромосом того или иного вида принято называть кариотипом. Кариотип характеризуется числом, размером, парностью и формой хромосом. Митоз, его биологическое значение. Стадии митоза. Митоз – непрямое деление клетки, наиболее распространенный способ репродукции эукариотических клеток. Профаза занимает 0,60 времени всего митоза, метафаза – 0,05, анафаза – 0,05 и телофаза – 0,3 времени всего митоза. Для большинства клеток млекопитающих фазы митоза длятся около 12 часов. В профазе две центриоли начинают расходиться к противоположным полюсам ядра. Ядерная мембрана разрушается; одновременно специальные белки объединяются, формируя микротрубочки в виде нитей. Центриоли, расположенные теперь на противоположных полюсах клетки, оказывают организующее воздействие на микротрубочки, которые в результате выстраиваются радиально, образуя структуру, напоминающую по внешнему виду цветок астры («звезда»). Другие нити из микротрубочек протягиваются от одной центриоли к другой, образуя веретено деления. В это время хромосомы спирализуются и вследствие этого утолщаются. Они хорошо видны в световом микроскопе, особенно после окрашивания. Считывание генетической информации с молекул ДНК становится невозможным: синтез РНК прекращается, ядрышко исчезает. В профазе хромосомы расщепляются, но хроматиды все еще остаются скрепленными попарно в зоне центромеры. Центромеры тоже оказывают организующее воздействие на нити веретена, которые теперь тянутся от центриоли к центромере и от нее к другой центриоли. В метафазе спирализация хромосом достигает максимума, и укороченные хромосомы устремляются к экватору клетки, располагаясь на равном расстоянии от полюсов. Образуется экваториальная, или метафазная, пластинка. На этой стадии митоза отчетливо видна структура хромосом, их легко сосчитать и изучить их индивидуальные особенности. В каждой хромосоме имеется область первичной перетяжки, к которой во время митоза присоединяются нить веретена деления и плечи. Хромосома состоит из двух хроматид, соединенных между собой только в области центромеры. В анафазе вязкость цитоплазмы уменьшается, центромеры разъединяются, и с этого момента хроматиды становятся самостоятельными хромосомами. Нити веретена деления, прикрепленные к центромерам, тянут хромосомы к полюсам клетки, а плечи хромосом при этом пассивно следуют за центромерой. Таким образом, в анафазе хроматиды удвоенных еще в интерфазе хромосом точно расходятся к полюсам клетки. В этот момент в клетке находятся два диплоидных набора хромосом (4n4с). В телофазе хромосомы раскручиваются, деспирализуются. Из мембранных структур цитоплазмы образуется ядерная оболочка. В это время восстанавливается ядрышко. На этом завершается деление ядра (кариокинез), затем происходит деление тела клетки (или цитокинез). При делении животных клеток на их поверхности в плоскости экватора появляется борозда, постепенно углубляющаяся и разделяющая клетку на две половины – дочерние клетки, в каждой их которых имеется по ядру. В результате митоза происходит точное распределение генетического материала между двумя дочерними клетками. Митоз обеспечивает поддержание постоянства числа хромосом в ряду поколений и служит клеточным механизмом процессов роста, развития организма, регенерации, бесполого размножения. При нарушении хода митоза, происходящего под действием некоторых факторов, наблюдается нерасхождение хромосом, нарушение их строения, повреждение веретена деления. Вследствие повреждений хромосом возникают различные мутации. Гомозигота_и_гетерозигота.__Наследственность'>Генотип и фенотип. Аллель. Гомозигота и гетерозигота. Наследственность – это способность живых организмов передавать потомкам морфологические, физиологические, биохимические признаки, особенности онтогенеза и обмена веществ. Благодаря наследственности обеспечивается материальная и функциональная преемственность в непрерывном ряде поколений. Внешним проявлением наследственности есть структурное и фенотипическое сходство родителей и потомков и всех особей, связанных родством. Генотип – это совокупность всех генов данной клетки, данного организма. Генотип состоит из хромотипа и плазмотипа. Проявление каждого гена зависит от других генов и от факторов внешней среды. Фенотип – совокупность признаков и свойств организма, которая формируется в онтогенезе в процессе взаимодействия генотипа и условий внешней среды. Признак – это любая особенность морфологии, физиологии, этологии, биохимии. Аллельные гены – это пара генов, определяющих контрастные признаки организма. Аллель – каждый ген аллельной пары. Альтернативные признаки – это взаимоисключающие, контрастные признаки (жёлтый – зелёный; высокий – низкий). Доминантный признак (преобладающий) – это признак, проявляющийся у гибридов первого поколения при скрещивании представителей чистых линий. Рецессивный признак (подавляемый) – это признак, не проявляющийся у гибридов первого поколения при скрещивании представителей чистых линий. Гомозигота – клетка или организм, содержащие одинаковые аллели одного и того же гена (АА или аа). Гетерозигота – клетка или организм, содержащие разные аллели одного и того же гена (Аа). Гибридологический метод – метод, который предполагает изучение признаков родительских форм, проявляющихся в ряду поколений у потомства, полученного путём гибридизации (скрещивания). Мутагенез. Классификация мутаций. Радиационный мутагенез. Зависимость частоты мутаций от вида облучения, мощности, дозы и чувствительности организма. Механизм возникновения мутаций. Процесс образования мутаций во времени и пространстве называется мутагенезом. Вещества, вызывающие мутации в клетках – мутагены. Общие закономерности мутагенеза Мутации возникают не мгновенно. Вначале возникает предмутационное состояние клетки. Репарационные системы стремятся устранить это состояние, и тогда мутация не реализуется. Однако при большом числе повреждений ДНК мутации могут стать необратимыми (не успевают исправлять повреждения, могут повреждаться сами ферменты систем репарации). Необратимые повреждения ДНК приводят к появлению мутаций. Мутации могут происходить под влиянием как внешних, так и внутренних воздействий. Генеративные и соматические; точечные, или генные, хромосомные, геномные мутации. 1901 г., де Фриз. Свойства: возникают внезапно, и мутировать может любая часть генотипа; чаще бывают рецессивными и реже – доминантными; могут быть вредными (большинство), нейтральными и полезными (очень редко) для организма; передаются из поколения в поколение; представляют собой стойкие изменения наследственного материала; могут повторяться. Точковые мутации – замена, выпадение или вставка одного или нескольких нуклеотидов. По эффекту бывают нонсенс-, сайленс- и миссенс-мутации, со сдвигом или без сдвига рамки считывания. Мутации могут вести к нарушению сплайсинга (сшивания экзонов и вырезания интронов). Отдельный тип мутаций – «динамические» мутации, которые характеризуются нарастанием триплетных повторов в гене. Хромосомные мутации – возникают из-за нарушения целостности хромосом. Характеризуются делецией, дупликацией или транслокацией участка хромосомы, а также транслокацией (обычной, робертсоновской, реципрокной), транспозицией и инверсией. Фрагмент хромосомы без центромеры, отделившийся при разрыве, может быть утрачен клеткой при очередном митозе. К геномным мутациям относят изменение числа геномов или числа отдельных хромосом: анеу-, поли- и гаплоидии. Радиационный мутагенез – возникновение мутаций под влиянием ионизирующих излучений и УФО. Различают спонтанный (естественный) мутагенез, происходящий под действием космической радиации или радиации, не контролируемой человеком (подземные радиоактивные элементы), и индуцированный (искусственный), осуществляемый в контролируемых человеком условиях; второй тип достаточно широко применяют в селекции (особенно в селекции микроорганизмов и растений). Повышение частоты вредных мутаций в результате увеличения содержания в биосфере радиоактивных изотопов – одна из основных опасностей радиоактивного загрязнения биосферы. Физические мутагены: ионизирующее излучение (α-, β-, γ-, нейтронное и рентгеновское излучение), коротковолновое ультрафиолетовое излучение, СВЧ-излучение, действие экстремальных температур. Действие ионизирующего излучения основано на ионизации компонентов цитоплазмы и ядерного матрикса. При ионизации возникают высокоактивные свободные радикалы, которые различным образом действуют на клеточные структуры. Непосредственное воздействие частиц с высокой энергией на ДНК, которое приводит к ее разрывам: одиночным (под воздействием γ-квантов, рентгеновских лучей) или множественных (под воздействием α-частиц, нейтронного излучения). Это универсальный механизм возникновения хромосомных перестроек на всех стадиях клеточного цикла, но он действует очень грубо – обычно клетки теряют способность к нормальному делению и погибают. К разрывам ДНК приводит и УФО. Наибольшей радиочувствительностью обладают клетки на стадии ранней профазы и во время синтетического периода интерфазы. Повреждение нуклеотидных оснований происходит значительно чаще, чем сахаро-фосфатного скелета. Пиримидиновые основания в 2 раза чувствительнее к облучению, чем пуриновые основания. Наиболее радиочувствителен тимин. Частота индуцированных генных мутаций пропорциональна дозе облучения, тогда как частота хромосомных аберраций под действием ионизирующего облучения возрастает пропорционально квадрату дозы. Чувствительность разных организмов к ионизирующему излучению различна: для человека 700 рентген – смертельная доза, для бактерий, вирусов таковой есть доза в миллионы рентген. Частота мутаций зависит и от обычных факторов среды. Так, повышение на 10°С температуры в местообитании вызывает 5-кратное увеличение частоты точечных мутаций. Мутагенез. Классификация мутаций. Химический мутагенез. Особенности мутагенного действия различных классов химических соединений. В настоящее время известно большое количество химических веществ, которые являются мутагенными, т. е. способными вызывать в ДНК разного рода нарушения. Среди химических веществ, обладающих специфической мутагенной активностью, выделяют соединения, мутагенные в отношении как реплицирующейся, так и нереплицирующейся ДНК (алкилирующие соединения, окислители-восстановители) и соединения, мутагенные только в отношении реплицирующейся ДНК. Также химические мутагены делят на мутагены прямого действия (соединения, реакционная способность которых достаточна для химической модификации ДНК, РНК и некоторых белков), и непрямого действия (промутагены – вещества, которые сами по себе инертны, но превращаются в организме в результате ферментативного окисления). Мишенью действия мутагенов в клетке являются ДНК и некоторые белки. Ряд мутагенов вызывают мутации, не связываясь ковалентно с ДНК. В этом случае матричный синтез на ДНК протекает с ошибками. В синтезируемой нити ДНК оказывается на один нуклеотид больше или меньше обычного и возникают мутации. Существуют мутагены, ингибирующие синтез предшественников ДНК. В результате происходит замедление или даже остановка синтеза ДНК. Алкилирующие агенты – это большая группа химических мутагенов, которые переносят алкильные группы на биополимеры. К ним относятся этиленимины, алкилалкансульфаты, эпоксиды, многоатомные спирты, серный и азотистый иприт, который непосредственно взаимодействует с ДНК. Алкилирующие соединения вызывают поперечные сшивки цепей в молекуле ДНК, приводящие к разрыву хромосом и появлению хромосомных аберраций. Под действием алкилирующих веществ могут образовываться сшивки ДНК-белок. Некоторые из алкилирующих агентов в природе не встречаются, их не распознают ферменты защитных систем. Такие вещества называются супермутагенами (например, N-метил-N-нитрозомочевина). Они применяются в селекции растений для получения индуцированных мутаций; их используют также как стимуляторы роста (в сверхмалых концентрациях); Соединения, мутагенные только в отношении реплицирующейся ДНК – к ним относятся производные пуринов и пиримидинов, акридиновые красители. Аналоги оснований, по молекулярной структуре похожие на основания, входящие в цепи ДНК, приводят к мутациям, так как они могут существовать в альтернативных (таутомерных) состояниях. Так, 5-бромурацил спаривается с аденином, в более редком таутомерном состоянии – с гуанином. Акридиновые красители (акридин оранжевый, профлавин и др.) индуцируют мутации типа сдвига рамки считывания. Они внедряются между соседними основаниями в ДНК. Молекула ДНК увеличивается на длину одного нуклеотида и при последующей репликации напротив внедряющейся молекулы красителя встраивается дополнительный дезоксирибонуклеотид, что и изменяет рамку считывания. Установлено, что мутагенной активностью обладает несколько тысяч химических соединений. Однако в отличие от ИО и УФО для химических мутагенов характерна специфичность действия, зависящая от природы объекта и стадии развития клетки. При взаимодействии химических мутагенов с компонентами наследственных структур (ДНК и белками) возникают первичные повреждения последних. В дальнейшем эти первичные повреждения ведут к возникновению хромосомных и геномных мутаций. Повреждающие агенты могут быть экзогенными (большинство известных мутагенов) или эндогенными (быть следствием ошибок метаболизма, нарушений функций генов системы детоксикации токсических метаболитов (свободные радикалы, перекисные соединения и т. д.) или генов репарации ДНК, результатом стресса или нарушений гормонального гомеостаза). В последнем случае особенно важную роль в механизмах нерасхождения хромосом отводят процессам внутрифолликулярного и внефолликулярного перезревания гамет, подробно исследованных в экспериментах на мышах и золотистых хомячках. Считается, что длительное пребывание яйцеклеток в фолликулах на фоне постоянно меняющегося гормонального фона, а также достаточно долгая (несколько дней) миграция овулировавшей яйцеклетки к месту оплодотворения приводят к кристаллизации и дегенеративным изменениям актиновых нитей митотического веретена и, как следствие, к нарушению сегрегации мейотических и митотических хромосом. В качестве мутагенных факторов рассматривают курение, различные химические, в том числе и лекарственные, препараты (антифолиевые препараты, цитостатики). Существует множество иных химических факторов, обладающих мутагенным, канцерогенным и тератогенным действием. Например, ионы тяжелых металлов, связываясь с ферментами репликации, репарации и рекомбинации, снижают их ферментативную активность. |