Генетика. Вопросы к экзамену. Генетика. Вопросы. Вопросы для сдачи экзаменов по дисциплине Общая и медицинская генетика
 Скачать 1.29 Mb. Скачать 1.29 Mb.
|

|
Вопросы для сдачи экзаменов по дисциплине «Общая и медицинская генетика» Этапы развития генетики, ее связь с другими науками. Вклад отечественных ученых в ее развитие. 1854-1863 гг. – исследования Менделя. 1866 г. – публикация его исследований, 2 выступления в Брюннском обществе. 1900 г. – повторное «открытие» законов Менделя Фризом, Корренсом, Чермаком, год рождения генетики. 1901 г. – де Фриз описывает мутации. 1902 г. – Бовари и Сэттон независимо пришли к выводу, что законы Менделя и поведение хромосом при митозе и мейозе соответствуют друг другу. Ученые предположили, что гены находятся в хромосомах. У. Сэттон высказал гипотезу о том, что гены, расположенные в одной хромосоме, будут наследоваться вместе. 1906 г. – английский ботаник Бетсон предложил название «генетика»; он и Пеннет наблюдали сцепление признаков, но не смогли объяснить и доказать хромосомную природу этого явления. 1911г. – Морган с сотрудниками доказал, что гены находятся в хромосоме и располагаются линейно. 1915 гг. – Морганом была сформулирована хромосомная теория наследственности. 1925 г. – Надсон и Филиппов доказали возможность получения искусственных мутаций у грибов под действием ионизирующего излучения. 1927 г. – Меллер показал, что мутации можно вызвать искусственно, воздействуя рентгеновскими лучами. 1928 г. – Кольцов выдвинул гипотезу о матричном синтезе и предположил, что гены являются белками (теория была дополнена в 1935 г.) 1940-ые гг. – Бидл и Тейтум выдвинули гипотезу «один ген – один фермент», которая впоследствии дополнялась. Доказанная цепочка «ген → фермент → реакция → признак» вдохновила большое количество исследователей на поиски генных дефектов, вызывающих различные заболевания. Возможность заменять «пораженные» гены «здоровыми» создала предпосылки для новой области науки – генной инженерии; Чаргафф показал, что общее количество аденина в каждой молекуле ДНК равно количеству тимина, а количество гуанина – количеству цитозина. Это явление было названо комплементарностью (правило Чаргаффа). 1950-е гг. – Уилкинс и Франклин получили рентгенограммы нитей ДНК. 1953 г. – на основании анализа данных Уилкинса и Чаргаффа Уотсон и Крик расшифровали химическую структуру молекулы ДНК. Открытие структуры нуклеиновых кислот позволило химикам синтезировать их искусственно «в пробирке». 1955 г. – Очоа впервые синтезировал РНК. 1957 г. – Корнберг впервые синтезировал ДНК, положено начало генной инженерии. 1958 г. – Крик сформулировал основной принцип передачи генетической информации «ДНК → РНК → белок» получивший название «центральная догма молекулярной биологии». 1958 г. – Мезельсон и Сталь установили полуконсервативный способ репликации ДНК. 1961 г. – Жакоб и Моно открыли теорию оперона. 1961-1966 гг. – Ниренберг, Корана и Очоа расшифровали генетический код. 1977 г. – открытие экзонов и интронов; метод секвенирования Сэнгера. Конец 70-ых – изучение транспозонов и мобильных элементов. 1995 г. – появился институт исследования генома человека, возглавляемый Ф. Коллинзом. Там проводилась программа «Геном человека» – основная задача которой заключалась в секвенировании всей ДНК, составляющей геном. 1997 г. – расшифровка генома E. coli, клонирование овечки Долли. Подобные опыты проводились и ранее: например, в 1940 г. русский эмбриолог Лопашев разработал метод пересадки ядер в яйцеклетку тритона. Правда, целью исследования было узнать, могут ли работать гены ядра дифференцированных клеток, помещенные в зиготу. 2000 г. – расшифровка генома дрозофилы. 2003 г. – расшифровка генома человека. Важнейшим итогом стало определение общего числа генов человека (около 25 000-30 000), большая часть которых (около 22 000) уже идентифицирована. Средний размер одного гена составляет около 3 тыс. п. н. Структура геномов людей разных национальностей, разных расовых и этнических групп на 99,9% идентична. Межиндивидуальная вариабельность даже при секвенировании генов представителей белой, желтой и черной рас не превышала 0,1 % и была обусловлена, главным образом, однонуклеотидными заменами. Н. П. Бочков – основатель кафедры мед. генетики в Сече, вплоть до 2009 ее глава. Исследовал радио- и химический мутагенез, занимался проблемой географического распределения наследственных заболеваний. Н. И. Вавилов – ввёл в употребление термин «селекция», лично занимался селекцией множества видов полезных культур. Собрал огромную коллекцию семян культурных растений, посвятил себя борьбе с голодом. По иронии судьбы во времена Сталинских репрессий и «лысенковщины» погиб от голода и был похоронен в Саратове в ящике из семян. Автор закона гомологических рядов, теории происхождения культурных растений, географического распределения генетических центров, генотипического иммунитета, ботанико-географических основ селекции, научных основ селекции пшеницы, внедрил новые более перспективные сорта пшеницы, исключив все неперспективные, продвигал растениеводство на Крайний Север, в зоны полупустынь, строил опытные станции по всей стране. Асанов Алий Юрьевич – профессор, ДМН, заведующий кафедрой медицинской генетики, 86 научных статей в области генетики. Автор учебника «Основы генетики и наследственных нарушений развития у детей». Структура ДНК. Комплементарность цепей. Репликация ДНК. Репарация ДНК. Дезоксирибонуклеиновая кислота (ДНК) – носитель наследственной информации у эукариот и прокариот, а рибонуклеиновая кислота (РНК) – может быть у вирусов. Прямые данные о способности ДНК передавать генетическую информацию впервые были получены в экспериментах Гриффитса (1928) с убитыми вирулентными и живыми невирулентными пневмококками. В опытах Эвери и Маккарти (1944) было показано, что фактор, отвечающий за трансформацию бактерий, разрушается ДНКазой и остаётся сохранным после действия протеаз. Впоследствии было показано, что именно нуклеиновые кислоты отвечают за хранение, передачу и реализацию наследственной информации. М 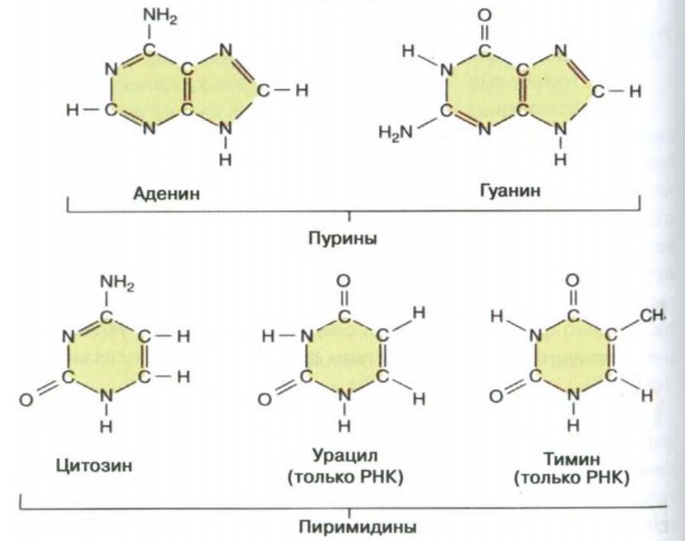 олекула ДНК – биополимер, состоит из сложных соединений – нуклеотидов, A-, B-, Z-формы, Z – левозакрученная, 12 п. н., A – 11, B – 3,4 нм и 10 п. н. за оборот. олекула ДНК – биополимер, состоит из сложных соединений – нуклеотидов, A-, B-, Z-формы, Z – левозакрученная, 12 п. н., A – 11, B – 3,4 нм и 10 п. н. за оборот.В-форма обычно находится в водных средах с небольшим количеством солей, А-форма – в обезвоженных средах и с большим количеством ионов, Z-форма существует только в присутствии особых белков, стабилизирующих её структуру. Азотистое основание (аденин, гуанин, цитозин, тимин) + дезокисирибоза (моносахарид) + фосфорный остаток. Биологический смысл – точная передача наследственной информации. Фермент ДНК-полимераза может двигаться вдоль цепи ДНК только в направлении 3’ → 5’. Количество ДНК постоянно и составляет 6,6 х 10-12 г. Функции ДНК: наследственная; структурная; определяет видоспецифичность. Комплементарность – это химическое и морфологическое соответствие нуклеотидов друг другу в парных цепочках ДНК. Правило Чаргаффа: количество пуринов равно количеству пиримидинов. Затем рентгеноструктурный анализ Уилкинса и Франклин, затем – модель ДНК Уотсона и Крика: две цепи, соединённые водородными связями, каркас составляют сахарные остатки и фосфорные, а основания находятся внутри, причём для двух пуринов места внутри бы не хватило, а пиримидины находились бы далековато друг от друга, поэтому напротив одного пурина находится пиримидин и наоборот (объяснение правила Чаргаффа). Водородные связи + гидрофобные стекинг-взаимодействия межплоскостными взаимодействия обеспечивают стабильность структуры. Репликация – это процесс самоудвоения ДНК, осуществляемый в ядре под контролем ферментов. Наиболее легко расплетаются участки, богатые А=Т связями. Топоизомеразы снижают напряжённость спирали, разрывая фосфодиэфирные связи и пропуская целую нить ДНК через разрыв, а затем сшивая его обратно. Праймаза способна «работать» с одноцепочечной ДНК в отсутствии 3’-OH-конца, а остальные полимеразы – нет, поэтому синтезируется затравка. У прокариот репликация начинается в ориджинах, у эукариот – в точках ARS (автономно реплицирующихся последовательностях). Деспирализация с помощью инициирующих белков на определенном участке, хеликаза, SSB-белки, праймаза, ДНК-полимераза (δ/ε), ДНК-полимераза β, ДНК-лигаза. Полуконсервативная модель. Репарация – исправление химических повреждений и разрывов в молекулах ДНК, повреждённой при нормальном биосинтезе ДНК или в результате воздействия физических или химических агентов. Ряд наследственных болезней (например, пигментная ксеродерма из-за УФ) связан с нарушениями систем репарации. Очень редко происходят повреждения, затрагивающие обе цепи ДНК. 1) Прямая репарация – наиболее простой путь устранения повреждений в ДНК, в котором обычно задействованы специфические ферменты, способные быстро (как правило, в одну стадию) устранять соответствующее повреждение. Пример: фотолиаза, которая, присоединяясь к пиримидиновому димеру, может расщепить его при поглощении энергии фотона. 2) Эксцизионная репарация – удаление повреждённых азотистых оснований из ДНК и последующее восстановление нормальной структуры молекулы, является наиболее распространенным способом репарации. N-гликозилазы (оксиметилурацил, гипоксантин, 5-метилурацил, 3-метиладенин, 7-метилгуанин и т.д.) отщепляют повреждённые нуклеиновые основания. Дальнейший процесс репарации может осуществляться с помощью ДНК-инсертазы, которая вставит в образовавшийся АП-сайт необходимый нуклеотид. Или с помощью ДНК-эндонуклеазы, которая найдёт искажение спирали и вырежет 8 нуклеотидов в направлении к 5’концу и 3-4 нуклеотида в направлении к 3’концу по обе стороны от АП-сайта, затем экзонуклеазы удалят и разрушат вырезанный участок ДНК, ДНК-полимераза β заполнит образовавшийся участок, а ДНК-лигаза соединит разрыв цепи. Может быть до-, пострепликативной или протекать прямо во время репликации. 3) SOS-репарация – в случае необходимости репликации ДНК, но при наличии в ней повреждений, которые не смогла устранить ни одна из вышеописанных систем, активизируется механизм данной репарации. Индуцируется синтез белка, который присоединяется к ДНК-полимеразному комплексу и позволяет строить дочернюю ДНК напротив дефектных звеньев матричной цепи. В результате ДНК оказывается удвоенной, хотя и с ошибкой, но это дает возможность провести клеточное деление. С дефектом системы репарации связаны некоторые наследственные болезни. Центральный постулат Ф. Крика. Генетический код, его свойства. Рибонуклеиновые кислоты, основные классы, участие в реализации экспрессии генов. ДНК – РНК – белок, 50-ые годы – постулат. Генетический код – это система записи наследственной информации в молекулах нуклеиновых кислот, основанная на определённом чередовании последовательностей нуклеотидов в ДНК или РНК. Свойства: триплетность, вырожденность, однозначность, полярность, неперекрываемость, компактность, универсальность. Триплет – наименьшая структурная единица. Кодон – наименьшая функциональная единица, кодирует одну аминокислоту или является стоп-кодоном. Стоп-кодонов три – УАА, УАГ, УГА. Полярность – считывание информации с ДНК и с мРНК происходит только в одном направлении. Структуры более высокого порядка определяются структурами более низкого порядка. Неперекрываемость – нуклеотид одного кодона не может быть одновременно нуклеотидом другого кодона. Перекрывающийся код найден у некоторых фагов. Компактность – между кодонами нет знаков препинания. Универсальность – код практически одинаков для всех живых организмов. Исключения из универсальности были обнаружены в митохондриях некоторых видов животных. Это касалось кодона терминатора УГА, который читался так же, как кодон УГГ, кодирующий триптофан. Были найдены и другие отклонения. Рибонуклеиновые кислоты – выполняют роль посредников в передаче информации от ДНК в цитоплазму. ДНК служит матрицей для синтеза РНК, которые являются матрицами синтеза белков, либо входят в состав рибосом, сплайсосом, либо переносят аминокислоты. РНК синтезируется в виде длинных неразветвленных полимерных молекул, которые способны образовывать двойные спирали, если различные части цепи имеют антипараллельные комплементарные сегменты. РНК некоторых вирусов является носителем наследственной информации вместо ДНК. м-РНК составляет 0,5-5% от всех видов РНК клетки, включает от 300 до 30 000 нуклеотидов. т-РНК составляет 10-15 % РНК клетки, состоит из 70-85 нуклеотидов. р-РНК составляет 80-85% РНК клетки, состоят из 3-5 тыс. н. Гетерогенная ядерная РНК (гяРНК) – смесь транскриптов многих ядерных генов, локализована в ядре. Некоторые из них являются первичными транскриптами и имеют такую же длину, как и гены, с которых они скопированы, другие – частично подверглись процессингу и сплайсингу и утратили ряд интронов, превратившись в зрелые мРНК. Малые ядерные РНК (мяРНК) – короткие стабильные молекулы, большинство которых в составе нуклеопротеидных частиц присутствуют в ядре. Они обнаружены в составе сплайсосом. Эти РНК называют U-PHK из-за необычайно большого содержания урацила и его модифицированных форм. Нуклеотидные последовательности всех U-PHK позвоночных совпадают на 95%. Малые ядрышковые РНК (мяоРНК) – вместе с нуклеазами принимают участие в процессинге рРНК. Локализованы в регионе ядрышка. Гены мяоРНК содержатся в интронах других генов, иногда единственной функцией последних служит производство мяоРНК. Малые цитоплазматические РНК (мцРНК) – известно, что большинство этих РНК находится в цитоплазме, участвует в РНК-интерференции, они аналогичны мяРНК. Малые интерферирующие РНК (миРНК) – получаются из протяженной двуцепочечной РНК, осуществляют деградацию мРНК (запуская РНК-интерференцию) и/или модификацию хроматина. МикроРНК (мкРНК) – образуются из крупных первичных транскриптов, которые имеют частично комплементарные области, образующие большое количество шпилек, участвуют в РНК-интерференции. piРНК – в комплексе с белками PIWI из семейства Argonaut участвуют в сайленсинге транспозонов, поддерживают стабильность зародышевой линии и, возможно, в эпигенетической наследственности. РНК-транскрипт сразу же по мере удлинения взаимодействует с белком, фиксируется химическими связями, одновременно в нём становятся доступными места модификации. РНК, связанная с белком, носит название рибонуклеопротеид (информосома). При выходе из ядра одни РНК остаются в соединении с белком, другие выходят из комплекса и принимают участие в трансляции. Метилазы метилируют собственный РНК-транскрипт в тех сайтах, которые могут быть разрезаны рестриктазами (защищают от трансляции с чужеродной ДНК/РНК). У эукариот РНК-транскрипт метилируется в меньшей степени. На растущей мРНК имеется специальная последовательность нуклеотидов (ААУААА). Поли-А-полимераза находит её, разрезает про-мРНК в этом месте и формирует полиадениловый хвостик. Поли-А последовательности также участвуют в терминации синтеза про-мРНК. РНК-полимераза, формируя последовательность ААУААА, получает сигнал о завершении синтеза РНК-транскрипта. Но синтез сразу не прекращается. Полная остановка его наступает после того, как РНК-полимераза встречает на матричной нити ДНК специфическую последовательность нуклеотидов (у разных генов она разная), которая даёт сигнал о прекращении синтеза РНК. Длина интронов может быть от 50 до 12000 нуклеотидов. Ген начинается и кончается экзоном. При мутации интрона сплайсинг может быть не закончен, и конченый продукт будет нести несвойственные последовательности нуклеотидов. Нарушение механизма сплайсинга – талассемии. Недостаток синтеза β-цепи гемоглобина – результат мутаций в двух участка гена – в сайте, ответственном за полиаденилирование, и в одном из интронов. В первом случае нарушается процесс формирования полиА-хвостика и формируется неполноценная β-цепь. Во втором случае сплайсосома не способна вырезать повреждённый интрон, и зрелая мРНК β-цепи гемоглобина не образуется. Ген. Структурные гены. Экзон-интронная структура гена эукариот, ее молекулярно-генетичеcкие доказательства. Уникальные и повторяющиеся последовательности. Ген – структурная и функциональная единица наследственности, контролирующая развитие определённого признака или свойства. Термин «ген» появился в 1909 г. В 1940 г., Бидл и Татум выдвинули гипотезу «1 ген – 1 фермент». Впоследствии она была дополнена до «1 ген – 1 белок». Сейчас считается, что 1 ген кодирует или 1 белок, или 1 нуклеиновую кислоту. Структурные гены кодируют клеточные белки с ферментативными или структурными функциями, РНК. Гены, определяющие место, время, длительность включения структурных генов – регуляторные гены. Из 25000 белков человека около 1% – структурные белки, 10% – ферменты, 90% – регуляторные белки. М 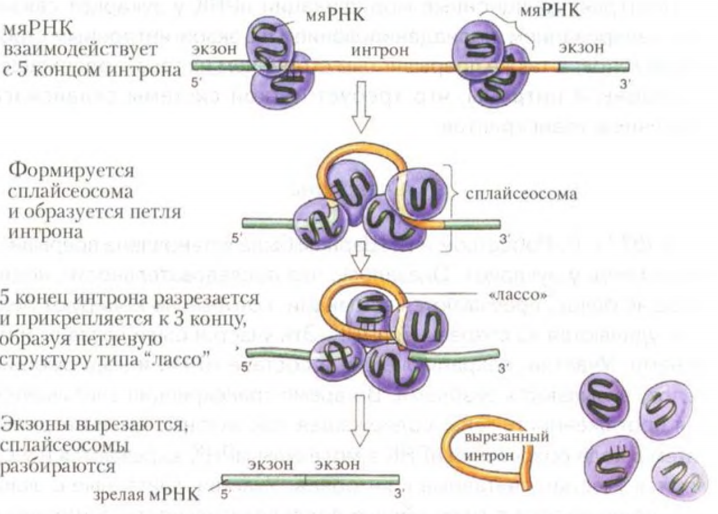 ногие эукариотические гены разделены на интроны и экзоны, прокариотические же гены состоят только из экзонов. Информационная РНК может содержать от одного до десятков интронов, размеры которых сильно варьируются. В число компонентов, катализирующих процесс сплайсинга, входят мяРНК и десятки белков, обладающих ферментативной активностью. Они образуют рибонуклеопротеидные частицы (мяРНП). Место соединения интрон/экзон узнается мяРНП, которые образуют более крупный комплекс, называемый сплайсосомой. ногие эукариотические гены разделены на интроны и экзоны, прокариотические же гены состоят только из экзонов. Информационная РНК может содержать от одного до десятков интронов, размеры которых сильно варьируются. В число компонентов, катализирующих процесс сплайсинга, входят мяРНК и десятки белков, обладающих ферментативной активностью. Они образуют рибонуклеопротеидные частицы (мяРНП). Место соединения интрон/экзон узнается мяРНП, которые образуют более крупный комплекс, называемый сплайсосомой.Механизм сплайсинга включает расщепление и прикрепление 5'-конца интрона к 3'-концу аденина сплайсосомы, при этом образуется петлевая структура, называемая «лассо». 3'-конец экзона затем обычно замещает 3'-конец интрона, два экзона соединяются вместе. Альтернативный сплайсинг позволяет организму синтезировать разные по структуре и свойствам белки на базе одного гена. В формировании альтернативных мРНК задействованы разные механизмы. Первый из них состоит в том, что для образования различных мРНК могут использоваться разные промоторы. В этом случае образуются транскрипты, имеющие разное количество экзонов. Такой механизм сплайсинга выявлен для иРНК легкой цепи миозина позвоночных, у коллагена – 2, пролактина – 3. Второй тип имеет место при изменении сайта полиаденилирования первичного транскрипта. Таким способом образуются два вида мРНК тяжелой цепи иммуноглобулинов. Третий тип включает вырезку различных экзонов из одинаковых иРНК. При этом для формирования зрелых РНК могут использоваться различные экзоны. Таким образом происходит сплайсинг иРНК тропонина Т скелетных мышц млекопитающих, содержащих 18 экзонов. При этом выбор экзонов зависит от стадии развития организма: экзон 16 присутствует в мРНК тропонина Т у взрослых, а экзон 17 – в мРНК этого белка у эмбрионов. РНК гена кальцитонина состоит из шести экзонов. В щитовидной железе вырезаются 5 и 6 экзоны и образуется кальцитонин. В гипоталамусе продуцируется белок CG RP пептид в результате удаления 4 и 6 экзонов. Последовательности ДНК, которые не повторяются, называются уникальной ДНК. Уникальные последовательности – это наиболее сложный компонент генома эукариот. У человека она составляет около 50% всей ДНК генома. Во фракцию уникальных последовательностей попадают многочисленные гены, кодирующие белки, интроны, размеры которых на порядок превышают размеры экзонов, содержащихся в гене, псевдогены – функционально неактивные «археологические» остатки прошлых генов. 1,5% – это экзоны и 5% – регуляторные последовательности. У человека повторы занимают по крайней мере 50% генома. Могут быть тандемными: они находятся на концах теломер и в области перетяжек. Теломеры состоят из небольших участков определенных последовательностей (по 68 нуклеотидных остатков), в которых многократно повторяются блоки ТТАГГГ. Синтезируются теломеразой. Из ТТАГГГ-блоков построены теломеры всех млекопитающих, рептилий, амфибий и рыб. В области первичных перетяжек располагается специфическая центромерная ДНК с часто повторяющимися короткими последовательностями ГЦ, АТ, с которыми взаимодействуют специальные белки кинетохоров, структур, обеспечивающих связь микротрубочек веретена деления с хромосомами. В группу тандемных повторов также входит рибосомная ДНК. |