Введение предмет, цели и задачи физиологии растений. Краткая история развития физиологии растений в мире, России, Казанском университете
 Скачать 0.49 Mb. Скачать 0.49 Mb.
|

|
Фотофосфорилирование: циклическое и нециклическое. Хемиосмотическая теория энергетического сопряжения Митчелла Сущность хемиосмотической теории:
На внутренней стороне мембраны тилакоида под действием света возбуждаются П680 и П700. Электроны от П680 захватываются акцепторами е- на наружной стороне и передаются на окисленную форму пластохинона (PQ) — липидорастворимого переносчика протонов и электронов, выполняющего функцию челнока. Принимая два электрона, 2PQ диффундируют к цитохромному комплексу и захватывают из стромы 2Н + . Электроны с 2PQH2 поступают в цепь: F.eSR -> цит. f -> Пц,-> П700, а ионы Н + попадают в полость тилакоида. Вторая пара ионов Н+ освобождается там же при фотоокислении воды. В ФС I электроны П700 захватываются акцептором А, передаются ферредоксину и затем НAДФ+ на наружной стороне мембраны, причем 1Н+ используется на восстановление НAДФ. Таким образом, из среды, окружающей тилакоид, при поглощении пигментами квантов света исчезают, а во внутренней полости тилакоида появляются протоны. В результате на мембране возникает электрохимический потенциал ионов Н + , который затем используется для фосфорилирования AДФ. Этот процесс называется нециклическим фотофосфорилированием. При циклическом фотофосфорилировании, когда функционирует только ФС I, электроны от Фд поступают на цитохромный комплекс с использованием пула PQ, который действует как переносчик электронов и протонов. Затем электроны через цитохром f и Пц возвращаются на основной энергетический уровень в П700, а протоны поступают в полость тилакоида
В результате фотохимических реакций в хлоропластах создается необходимый уровень АТР и NADPH. Эти конечные продукты световой фазы фотосинтеза стоят на входе в темновую фазу, где С02 восстанавливается до углевода. Химизм реакции цикла Кальвина Этот способ ассимиляции С02, присущий всем растениям. Фотосинтезирующие водоросли помещали в среду, содержащую 14С02, на разные промежутки времени, затем клетки быстро фиксировали, экстрагировали из них спирторастворимые вещества и определяли содержание 14С в различных соединениях после их разделения с помощью хроматографии. Оказалось, что после экспозиции в течение 1 мин 14С включался в С3 —С7-сахара и фосфосахара, в органические кислоты (яблочную, щавелевоуксусную, ФЕП), в аминокислоты (аланин, аспарагиновую кислоту). Если же время экспозиции было сокращено до 0,1—2 с, то большая часть метки обнаруживалась в фосфоглицериновой кислоте, в ее карбоксильной группе. Следовательно, 3-фосфоглицериновая кислота (ФГК) является первичным продуктом фотосинтеза. Природа первичного акцептора С02. Сначала предположили, что таким акцептором является какое-либо двухуглеродное соединение. Однако введение в инкубационную среду винилфосфата, фосфогликольальдегида и других веществ с С2 не приводило к увеличению содержания радиоактивной метки из С02 в ФГК. Тогда схему опыта видоизменили следующим образом. Водоросли экспонировали на свету при высокой (1 %-ной) концентрации С02, затем резко снижали его концентрацию до 0,003%. Расчет был на то, что в условиях дефицита С02 быстро накопится именно то соединение, которое служит акцептором С02. С помощью двумерной хроматографии удалось установить, что при отсутствии возможности карбоксилирования в клетках кратковременно возрастает концентрация рибулозо-1,5-дифосфата (рибулозо-1,5-бисфосфата). Отсюда возникло предположение, что первичная фиксация С02 происходит следующим образом : С5 + С02-> С6-► 2С3. Для проверки этой гипотезы в бесклеточный экстракт из листьев шпината или клеток хлореллы вносили меченный по 32Р рибулозо-1,5-дифосфат. На свету в экстракте появлялась радиоактивная ФГК. На основании полученных данных процесс первичной фиксации С02 можно записать следующим образом: 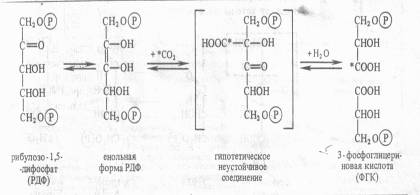 Эта реакция катализируется рибулозодифосфаткарбоксилазой (рибулозобисфосфаткарбоксилазой; другие названия — белок фракции I, карбоксидисмутаза). Максимум активности фермент имеет при рН 7,8 — 8,0 и нуждается в присутствии ионов Mg2 + . Дальнейшая работа в лаборатории Кальвина и в других лабораториях привела к расшифровке всех последующих реакций С3-пути фотосинтеза, который получил название цикла Кальвина. Этот цикл, весьма напоминающий обращенный пентозофосфатный путь дыхания, состоит из трех этапов: карбоксилирования, восстановления и регенерации. 1. Карбоксилирование. Молекулы рибулозо-5-фосфата фосфорилируются с участием АТР и фосфорибулозокиназы, в результате чего образуются молекулы рибулозо-1,5-дифосфата, к которым в свою очередь присоединяется С02 с помощью рибулозодифосфаткарбоксилазы. Полученный продукт расщепляется на две триозы: 2 молекулы 3-фосфо-глицериновой кислоты (3-ФГК). 2. Фаза восстановления. 3-ФГК восстанавливается до 3-фосфоглицеринового альдегида (3-ФГА) в два этапа. Сначала происходит фосфорилирование 3-ФГК при участии АТФ и фосфоглицераткиназы до 1,3-дифосфоглицериновой кислоты, а затем восстановление 1,3-ФГК с помощью НАДФH и дегидрогеназы фосфоглицеринового альдегида. 3. Фаза регенерации первичного акцептора диоксида углерода и синтеза конечного продукта фотосинтеза. В результате описанных выше реакций при фиксации трех молекул С02 и образовании шести молекул восстановленных 3-фосфотриоз пять из них используются затем для регенерации рибулозо-5-фосфата, а один — для синтеза глюкозы. 3-ФГА под действием триозофосфатизомеразы изомеризуется в фосфодиоксиацетон. При участии альдолазы 3-ФГА и фосфодиоксиацетон конденсируются с образованием фруктозо-1,6-дифосфата, у которого отщепляется один фосфат с помощью фруктозо-1,6-дифосфатазы. В дальнейших реакциях, связанных с регенерацией первичного акцептора С02, последовательно принимают участие транскетолаза и альдолаза. Транскетолаза катализирует перенос содержащего два углерода гликолевого альдегида от кетозы на адьдозу: 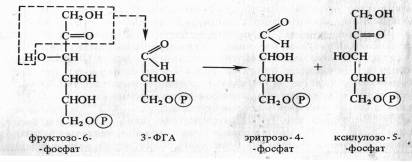 Альдолаза затем осуществляет перенос трехуглеродного остатка фосфодиоксиацетона на альдозу, в данном случае эритрозо-4-фосфат, в результате чего синтезируется седо-гептулозо-1,7-дифосфат. Последний дефосфорилируется и под действием транскетолазы из него и 3-ФГА образуются ксилулозо-5-фосфат и рибозо-5-фосфат. Две молекулы ксилулозо-5-фосфата при участии рибулозофосфатэпимеразы и одна молекула рибозо-5-фосфата с участием рибозофосфатизомеразы превращаются в три молекулы рибулозо-5-фосфата, с которого начинается новый цикл фиксации С02. Из оставшейся неиспользованной 6-ой молекулы 3-ФГА под действием альдолазы синтезируется (при повторении цикла) молекула фруктозо-1,6-дифосфата, из которой могут образовываться глюкоза, сахароза или крахмал: Таким образом, для синтеза одной молекулы глюкозы в цикле Кальвина необходимы 12 NADPH и 18 АТР, которые поставляются в результате фотохимических реакций фотосинтеза.
В работах Л. А. Незговоровой (1956—1957 гг.), было установлено, что при коротких экспозициях листьев кукурузы на свету 14С из 14С02 обнаруживается в аспарагиновой кислоте. В дальнейших исследованиях как советских, так и зарубежных специалистов эти представления были развиты, что привело к открытию C4-nymu углерода в фотосинтезе. Так, в 1960 г. Ю. С. Карпилов, а в 1963 г. И. А. Тарчевский и Ю. С. Карпилов представили данные о раннем образовании яблочной кислоты в листьях кукурузы. Г. П. Корчак и др. впервые показали, что дикарбоновые кислоты (яблочная и аспарагиновая) являются первичными продуктами фиксации С02 у сахарного тростника. Затем эти соединения через 3-ФГК и гексозофосфаты превращаются в сахара. Как новый тип фиксации С02, принципиально отличающийся от цикла Кальвина, этот цикл впервые описали австралийские ученые М. Д. Хетч и, К. Р. Слэк (1966). К группе растений с С4-путем фотосинтеза относятся сахарный тростник, кукуруза, сорго и др. Листья этих растений содержат два разных типа хлоропластов: хлоропласты обычного вида — в клетках мезофилла и большое количество крупных хлоропластов, часто не имеющих гран, — в клетках, окружающих проводящие пучки (обкладка). С02, диффундирующий в лист через устьица, попадает в цитоплазму клеток мезофилла, где при участии ФЕП-карбоксилазы вступает в реакцию с ФЕП, образуя щавелевоуксусную кислоту (оксалоацетат). Затем уже в хлоропластах оксалоацетат восстанавливается до яблочной кислоты (малата) за счет NADPH, образующего в ходе световой фазы фотосинтеза. Щавелевоуксусная кислота в присутствии NH4+ может превращаться также в аспартат. Затем малат (или аспартат) переносится в хлоропласты клетки обкладки сосудистого пучка, где он декарбоксилируется малик-энзимом (малатдегидрогеназой декарбоксилирующей) до пирувата и С02. В хлоропластах обкладки отсутствуют граны, а следовательно, слабо представлена ФС II, необходимая для нециклического транспорта электронов, однако в них в изобилии накапливается крахмал. Объясняется это тем, что в хлоропластах обкладки используется поставляемый малик-энзимом НAДФH, а также тот С02, который образовался при окислительном декарбоксилировании малата (или аспартата). В этих хлоропластах в процессе циклического фотофосфорилирования синтезируется большое количество АТР и фиксация С02 осуществляется по типу цикла Кальвина. У некоторых растений с С4-путем фотосинтеза (амарант, лебеда) яблочная кислота декарбоксилируется в митохондриях клеток обкладки с восстановлением NAD. Возникающий при расщеплении малата в хлоропластах клеток обкладки пируват перемещается назад в хлоропласты клеток мезофилла, где может снова превращаться в первичный акцептор С02 — ФЕП. Такая компартментация процессов позволяет растениям с С4-путем осуществлять фотосинтез даже при закрытых устьицах, так как хлоропласты клеток обкладки используют малат (аспартат), образовавшийся ранее, как донор С02. С4-растения могут также использовать С02, возникающий при фотодыхании. Закрывание устьичных отверстий в наиболее жаркое время дня сокращает потери воды за счет испарения (транспирации). Не удивительно поэтому, что к С4-растениям относятся многие виды засушливой тропической зоны. Растения с С4-путем фотосинтеза, как правило, устойчивы к засолению. Эффективность использования воды, т. е. отношение массы ассимилированного С02 к массе воды, израсходованной при транспирации, у С4-растений зачастую вдвое выше, чем у С3-растений. Таким образом, С4-растения имеют преимущества перед С3-растениями в засушливых местах обитания благодаря высокой интенсивности фотосинтеза даже при закрытых устьицах. Фиксация С02 с участием ФЕП и образование малата (аспартата) служит как бы насосом для поставки С02 в хлоропласты обкладки, функционирующие по С3-пути. САМ-тип метаболизма. Потоки метаболитов в хлоропласт и из него Суккуленты (роды Crassula, Bryophyllum и др.) также приспособились осуществлять фотосинтез в условиях резко засушливого климата. Для них характерен суточный цикл метаболизма С4-кислот с образованием яблочной кислоты ночью. В соответствии с английским выражением Crassulacean Acid Metabolism (САМ) этот тип фотосинтеза часто сокращенно называют САМ-метаболизм. Устьица этих растений днем обычно закрыты, что предотвращает потерю воды, и открываются ночью. С02 поступает в листья, где при участии содержащейся в цитоплазме ФЕП-карбоксилазы взаимодействует с фосфоенолпируватом, образуя оксалоацетат. Источник ФЕП-карбоксилазы взаимодействует с фосфоенолпируватом, образуя оксалоацетат. Источником ФЕП служит крахмал. То же самое происходит и с С02, который освобождается в клетках в процессе дыхания. Образовавшийся оксалоацетат восстанавливается под действием NADH-зависимой малатдегидрогеназы до яблочной кислоты, которая накапливается в вакуолях клеток листа. Это приводит к закислению клеточного сока в ночное время. Как и у С4-растений, оксалоацетат может быть источником аспартата, однако этот путь здесь менее выражен. Днем в условиях высокой температуры, когда устьица закрыты, малат транспортируется из вакуолей в цитоплазму и там декарбоксилируется при участии малатдегидрогеназы декарбоксилирующей (малик-энзима) с образованием С02 и пирувата. С02 поступает в хлоропласты и включается в них в цикл Кальвина, участвуя в синтезе сахаров. Таким образом, у растений с фотосинтезом по типу толстянковых много общего с С4-путем фотосинтеза. Однако при САМ-метаболизме фиксация С02 с образованием малата (ночью) и декарбоксилирование малата с высвобождением С02 и пирувата (днем) разделены во времени. У С4-растений эти же реакции разграничены в пространстве: первая протекает в хлоропластах мезофилла, вторая — в клетках обкладки. При достаточном количестве воды ряд растений с метаболизмом по типу толстянковых могут вести себя как С3-растения. В свою очередь некоторые растения с С3-путем фотосинтеза при недостатке воды проявляют черты САМ-метаболизма.
Исследования показали, что в растениях, в которых процесс фотосинтеза протекает по С4-пути, имеются два типа клеток и хлоропластов: 1) мелкие гранальные пластиды в клетках мезофилла листа; 2) крупные пластиды, часто лишенные гран, в клетках обкладки, окружающих сосудистые пучки. Клетки обкладки имеют утолщенные клеточные стенки, содержат большое количество хлоропластов и митохондрий, расположены вокруг сосудистых пучков в 1 или 2 слоя. У таких растений специфичное анатомическое строение листа. У них два типа фотосинтезирующих клеток: клетки обкладки, радиально покрывающие проводящие пучки в листе, и клетки мезофилла столбчатой и губчатой ткани. Это может служить диагностическим признаком на определение С4-растений. В структуре этих клеток также есть различия: в клетках обкладках в хлоропластах отсутствуют граны, они агранальные, а в клетках мезофилла все хлоропласты гранальные К группе с С4-путем фотосинтеза относятся сахарный тростник, кукуруза, сорго и др. Эти растения устойчивы к засолению. Эффективность использования воды , т.е. отношение массы ассимилированного СО2 к массе воды, израсходованной при транспирации, у С4 растений зачастую вдвое выше, чем у С3-растений. Таким образом, С4-растения имеют преимущества перед С3-растениями в засушливых местах обитания, благодаря высокой интенсивности фотосинтеза даже при закрытых устьицах. Фиксация СО2 с участием ФЕП и образование малата(аспартата) служит как бы насосом для поставки СО2 в хлоропласты обкладки, функционирующие по С3 пути.
Путь восстановления СО2 при фотосинтезе М. Кальвин изучал при относительно высоких концентрациях углекислого газа. При более низких концентрациях СО2, приближающихся к атмосферным, в хлоропласте образуется значительное количество фосфогликолевой кислоты. Это связано с тем, что в условиях низкого парциального давления СО2 и высокого парциального давления О2 РДФ-карбоксилаза может присоединять к рибулозо-1,5-дифосфату кислород; это значит, что она может выполнять не только карбоксилирующую, но и оксигенерирующую функции. Присоединение кислорода к молекуле рибулозодифосфата приводит к тому, что вместо двух молекул ФГК образуется одна молекула ФГК и одна молекула фосфогликолата. Последняя во время транспорта через мембрану хлоропласта дефосфорилируется при участии фосфогликолатфосфатазы и превращается в гликолат, который поступает в специальную органеллу-периксисому. В последней содержится каталаза – фермент, разлагающий перекиси. В периксисоме гликолат вступает в реакцию с кислородом воздуха, в результате образуется глиоксилат и перекись водорода. Реакцию катализирует гликолатоксидаза. Перекись сразу разрушается каталазой на воду и кислород, а глиоксилат в результате реакции трансаминирования, которая проходит при участии глиоксилатглицинаминотрансферазы, превращается в глицин, который перемещается в митохондрию. Тут из двух молекул глицина при участии серинглиоксилатаминотрансфераза синтезируется одна молекула серина, образуется СО2, аммиак и восстанавливается НАД+. СО2 или выделяется в атмосферу, это значит теряется для растения, или вновь может связаться в С3-цикле. Предполагают, что образующийся НАДН может быть связан с электрон-транспортной цепью дыхания, что приводит к синтезу АТФ при его окислении. Серин из митохондрий транспортируется вновь в периксисому, где после дезаминирования превращается в оксипируват, потом в глицерат, и наконец, в ФГК. ФГК – конечный продукт этого цикла может поступать в С3-цикл. Цикл превращения гликолата в триозофосфат(ФГК) называется гликолатным. Образовавшийся аммиак не выделяется в окружающую среду, а связывается в виде глутамина. Так как образуются кислоты, содержащие 2 атома углерода, то этот цикл получил название С2-цикла. С другой стороны, так как во время этих превращений происходит поглощение кислорода и выделение СО2, то этот процесс получил название фотодыхания. Фотодыхание (или световое дыхание) – это индуцированное светом поглощение О2 и выделение СО2, которое наблюдается в зеленых клетках. Интенсивность фотодыхания увеличивается при повышении освещенности. Хотя органеллой фотодыхания считается периксисома, в этом цикле непосредственное участие принимают хлоропласты и митохондрии. Интенсивное фотодыхание может снизить продуктивность С3-растений на 30 %, так как при фотодыхании происходит не поглощение СО2, а его потери. Из каждых 10 молекул риболозо-1,5-дифосфата, образовавшегося в С3-цикле и включившихся в фотодыхание, одна разрушается: 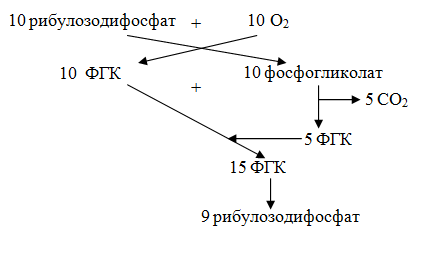 Низкую интенсивность фотодыхания в С4-растениях объясняют тем, что в них высокая концентрация СО2 в клетках-обкладки. В этих растениях РДФ-карбоксилаза находится только в клетках-обкладки, где и происходит процесс. Фотодыхание рассматривают как одну из необходимых функций зеленого растения. Во-первых, фотодыхание, как и С3-цикл, САМ-цикл и другие процессы, является источником метаболитов для разных синтезов. Во-вторых, что при образовании серина и глицина восстанавливается митохондриальная НАД+. В-третьих, выявлена связь фотодыхания с ассимиляцией азота. И, наконец, появилась теория, в соответствии с которой излишки продуктов цикла Кальвина служат сигналом, который увеличивает фотосинтез. Таким образом, соотношение фотосинтеза и фотодыхания выполняют регуляторную роль в растениях. |