Введение предмет, цели и задачи физиологии растений. Краткая история развития физиологии растений в мире, России, Казанском университете
 Скачать 0.49 Mb. Скачать 0.49 Mb.
|

Влияние внешних и внутренних факторов на интенсивность фотосинтезаВ физиологии растений пользуются двумя понятиями: истинный и наблюдаемый фотосинтез. Это обусловлено следующими соображениями. Скорость или интенсивность фотосинтеза характеризуется количеством СО2, поглощенного единицей поверхности листа в единицу времени. Определение интенсивности фотосинтеза проводят газометрическим методом по изменению (уменьшению) количества СО2 в замкнутой камере с листом. Однако, вместе с фотосинтезом идет процесс дыхания, во время которого выделяется СО2. Поэтому получаемые результаты дают представление об интенсивности наблюдаемого фотосинтеза. Для получения величины истинного фотосинтеза необходимо сделать поправку на дыхание. Поэтому перед опытом определяют в темноте интенсивность дыхания, а потом уже интенсивность наблюдаемого фотосинтеза. Затем количество СО2, выделенного при дыхании, прибавляют к количеству СО2, поглощенного на свету. Внося эту поправку, считают, что интенсивность дыхания на свету и в темноте одинакова. Но эти поправки не могут дать оценку истинного фотосинтеза потому, что, во-первых, при затемнении листа исключается не только истинный фотосинтез, но и фотодыхание; во-вторых, так называемое темновое дыхание в действительности зависит от света (см. дальше). Поэтому во всех экспериментальных работах по фотосинтетическому газообмену листа отдают преимущество данным по наблюдаемому фотосинтезу. Более точный метод изучения интенсивности фотосинтеза – метод меченных атомов (измеряют количество поглощенного 14СО2). В том случае, когда пересчет количества поглощенного СО2 на единицу поверхности трудно провести (хвойные, семена, плоды, стебель), полученные данные относят к единице массы. Учитывая, что фотосинтетический коэффициент (отношение объема выделенного кислорода к объему поглощенного СО2 равен единице, скорость наблюдаемого фотосинтеза можно оценивать по количеству миллилитров кислорода, выделенной единицей площади листа за 1 час. Для характеристики фотосинтеза пользуются и другими показателями: квантовый расход, квантовый выход фотосинтеза, ассимиляционное число. Квантовый расход – это отношение количества поглощенных квантов к количеству ассимилированных молекул СО2. Обратная величина названа квантовым выходом. Ассимиляционное число – это соотношение между количеством СО2 и количеством хлорофилла, который содержится в листе. Скорость (интенсивность) фотосинтеза – один из важнейших факторов, влияющих на продуктивность с/х культур, а значит и на урожай. Поэтому выяснение факторов, от которых зависит фотосинтез, должно вести к усовершенствованию агротехнических мероприятий. Теоретически скорость фотосинтеза, как и скорость любого многостадийного биохимического процесса, должна лимитироваться скоростью самой медленной реакции. Так, например, для темновых реакций фотосинтеза нужны НАДФН и АТФ, поэтому темновые реакции зависят от световых реакций. При слабой освещенности скорость образования этих веществ слишком мала, чтобы обеспечить максимальную скорость темновых реакций, поэтому свет будет лимитирующим фактором.Принцип лимитирующих факторов можно сформулировать следующим образом: при одновременном влиянии нескольких факторов скорость химического процесса лимитируется тем фактором, который ближе всех к минимальному уровню (изменение именно этого фактора будет непосредственно влиять на данный процесс). Этот принцип впервые был установлен Ф. Блекманом в 1915 г. С тех пор было неоднократно показано, что разные факторы, например концентрация СО2 и освещенность, могут взаимодействовать между собой и лимитировать процесс, хотя часто один из них все же главенствует. Освещенность, концентрация СО2 и температура – вот те главные внешние факторы, влияющие на скорость фотосинтеза. Однако большое значение имеет также водный режим, минеральное питание и др. Свет.При оценке действия света на тот или иной процесс важно различать влияние его интенсивности, качества (спектрального состава) и времени экспозиции на свету.При низкой освещенности скорость фотосинтеза пропорциональна интенсивности света. Постепенно лимитирующими становятся другие факторы, и увеличение скорости замедляется. В ясный летний день освещенность составляет примерно 100 000 лк, а для светового насыщения фотосинтеза хватает 10 000 лк. Поэтому свет обычно может быть важным лимитирующим фактором в условиях затенения. При очень большой интенсивности света иногда начинается обесцвечивание хлорофилла, и это замедляет фотосинтез; однако в природе, растения находящиеся в таких условиях, обычно тем или иным образом защищены от этого (толстая кутикула, опущенные листья и т. п.). Зависимость интенсивности фотосинтеза от освещенности описывается кривой, которая получила название световой кривой фотосинтеза (рис. 2.26). 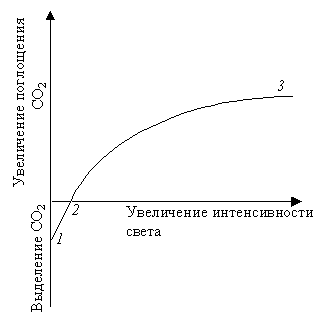 Рис. 2.26.Зависимость интенсивности фотосинтеза от освещенности (световая кривая фотосинтеза): 1 – скорость выделения СО2 в темноте (скорость дыхания);2 – компенсационная точка фотосинтеза; 3 – положение светового насыщения При слабом освещении в процессе дыхания выделяется больше СО2, чем связывается его в процессе фотосинтеза, поэтому начало световой кривой с осью абсцисс – компенсационная точкафотосинтеза, которая показывает, что в этом случае при фотосинтезе используется ровно столько СО2, сколько его выделяется при дыхании. Иными словами, со временем наступает такой момент, когда фотосинтез и дыхание будут точно уравновешивать друг друга, так что видимый обмен кислорода и СО2 прекратиться. Световая точка компенсации – это такая интенсивность света, при которой суммарный газообмен равен нулю. Световые кривые одинаковы не для всех растений. У растений, которые растут на открытых солнечных местах, поглощение СО2 увеличивается до тех пор, пока интенсивность света не будет равна полному солнечному освещению. У растений, которые растут на затененных местах (например, кислица), поглощение СО2 увеличивается только при малой интенсивности света. Все растения по отношению к интенсивности света делят на световые и теневые, или светолюбивые и теневыносливые. Большинство с/х растений является светолюбивыми. У теневыносливых растений, во-первых, световое насыщение происходит при более слабом освещении, во-вторых, в них компенсационная точка фотосинтеза наступает раньше, т. е. при меньшей освещенности (рис. 2.27). 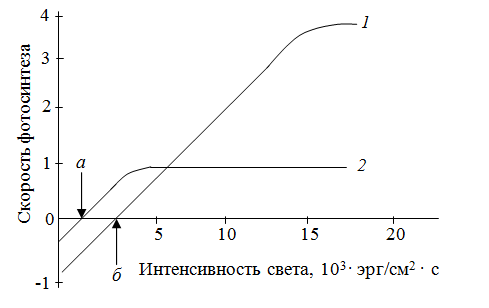 Рис. 2.27. Световые кривые светолюбивого (1) и теневыносливого (2) растения (а, б – компенсационные точки) Последнее связано с тем, что теневыносливые растения отличаются малой интенсивностью дыхания. В условиях слабой освещенности интенсивность фотосинтеза выше у теневыносливых растений, а при сильном свете, наоборот, – у светолюбивых. Интенсивность света влияет и на химический состав конечных продуктов фотосинтеза. Чем выше освещенность, тем больше образуется углеводов; при низкой освещенности – больше органических кислот. Опыты в лабораторных условиях показали, что на качество продуктов фотосинтеза влияет и резкий переход «темнота – свет» и наоборот. Сначала после включения света высокой интенсивности преимущественно образуются неуглеводные продукты из-за недостатка НАДФН и АТФ, и только через некоторое время начинают образовываться углеводы. И наоборот, после выключения света листья не сразу теряют способность к фотосинтезу, потому что на протяжении нескольких минут в клетках остается запас АТФ и НАДФ. После выключения света сначала тормозится синтез углеводов и только потом органических веществ и аминокислот. Основная причина этого явления обусловлена тем, что торможение превращения ФГК в ФГА (и через него в углеводы) происходит раньше, чем торможение ФГК в ФЕП (и через него в аланин, малат и аспарат). На соотношение образующих продуктов фотосинтеза влияет и спектральный состав света. Под влиянием синего света в растениях увеличивается синтез малата, аспартата и других аминокислот и белков. Эта реакция на синий свет выявлена и в С3-растениях и в С4-растениях. Спектральный состав света влияет и на интенсивность фотосинтеза (рис. 2.28). 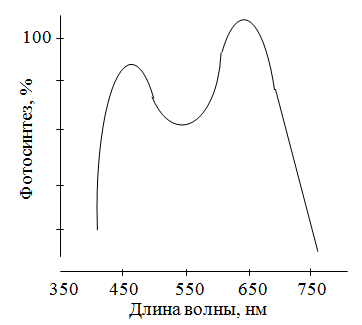 Рис. 2.28.Спектр действия фотосинтеза листьев пшеницы Спектр действия – это зависимость эффективности химического (биологического) действия света от длины его волны. Интенсивность фотосинтеза в разных участках спектра неодинакова. Максимальная интенсивность наблюдается при освещении растений теми лучами, которые максимально поглощаются хлорофиллами и другими пигментами. Интенсивность фотосинтеза наиболее высокая в красных лучах, потому что она пропорциональна не количеству энергии, а количеству квантов. Из суммарного уравнения фотосинтеза: 6СО2 + 6Н2О → С6Н12О6 + 6О2 следует, что для образования 1 моля глюкозы нужно 686 ккал; это значит, что для ассимиляции 1 моля СО2 нужно 686: 6 = 114 ккал. Запас энергии 1 кванта красного света (700 нм) равен 41 ккал/энштейн, а синего (400 нм) 65 ккал/энштейн. Минимальный квантовый расход при освещении красным светом равен 114: 41 ≈ 3, а в действительности тратиться 8–10 квантов. Таким образом, эффективность использования красного света 114/41 · 8 = 34 %, а синего 114/65 ·8 = 22 %. Изменение интенсивности дыхания в онтогенезе. Наиболее высокой обладают молодые органы и ткани растений, находящиеся в состоянии активного роста. Повышение интенсивности дыхания прекращается, когда заканчивается рост листа. Затем дыхание снижается до уровня, приблизительно равного половине максимума, и довольно долго остается без изменений. Цветение и плодоношение сопровождается усилением дыхания развивающихся цветков и плодов, что связано с образованием новых органов и тканей, обладающих высоким уровнем обмена веществ. Подъем интенсивности дыхания перед полным созреванием плодов называется климактерическим подъемом дыхания. Сходное явление наблюдается и при пожелтение листьев. Перед климактерическим подъемом дыхания в тканях резко усиливается образование этилена, который оказывает на обмен двоякое влияние. 1. Увеличивается проницаемость мембраны и усиливается гидролиз белков, в результате чего возрастает количество доступных дыханию субстратов. 2. В период подъема климактерического дыхания стимулируется синтез белков возможно дыхательных ферментов.  МИНЕРАЛЬНОЕ ПИТАНИЕ
Первые труды появляются в средние века. Среди них назовем «Научный трактат о различных почвах (солях) и сельском хозяйстве» французского естествоиспытателя Палисси. В этом сочинении впервые почва рассматривается как источник питания растений минеральными солями, высказывается мысль о необходимости возврата в почву минеральных веществ в виде удобрений. Ван-Гельмонт. Им был поставлен эксперимент с растениями, на основании которого был сделан ошибочный вывод о питании растений водой. Ива и горшок с землей (Водная теория, была ошибочна) Глаубер выдвинул гипотезу, согласно которой основой роста растений является селитра( «соль плодородия»), получаемую из смеси навоза с землей. Глаубер предположил, что она образуется из пищи животных. Он советовал вносить селитру под виноградники, смачивать раствором селитры высеваемое зерно. Лавуазье установил закон сохранения вещества, определил состав воздуха и процесс образования углекислого газа, сделал ряд других важных открытий. Вместе с тем он занимался и вопросами, относящимися к агрономической химии. Он писал: «Растения черпают материалы, необходимые для своей организации, в воздухе, который их окружает, в воде, вообще в минеральном царстве.». Пристли проделал замечательный опыт. Он взял стеклянный колпак, изолировал под ним мышь и горящую свечу, потом поместил мяту. Вывод: под действием зеленых растений воздух становится пригодным для дыхания и горения. Ингенгаус обнаружил, что только растения, причем только на свету, поглощают из воздуха углекислоту, а выделяют при этом кислород. Сами растения непрерывно дышат, но на свету значительно больше выделяют кислорода, а при отсутствии света сами используют некоторую часть O2. Сенебье и Соссюр представили экспериментальные доказательства минерального корневого и воздушного питания растений. Ученые показали, что в усвоении углекислого газа участвует «зеленый крахмал», то есть хлорофилл. Ученые показали, что под влиянием солнечной радиации листья испаряют влагу, способствуя этим доступу новых порций «соков» из почвы и корней в надземные органы растений. Под действием света они поглощают углекислый газ и выделяют кислород. Корни из почвы доставляют растениям лишь малую часть пищи, но эти минеральные вещества им совершенно необходимы. Тэер был сторонником гумусовой теории питания растений. Согласно этой теории, перегной непосредственно поглощается корнями и служит основной пищей растениям. Минеральные вещества эта теория ошибочно рассматривала лишь как вспомогательные, содействующие усвоению гумуса. Либих показал, что все минеральные соединения растения поглощают из почвы. Поэтому для восстановления почвенного плодородия эти вещества в почву необходимо возвращать, «закон возврата»; «закон минимума»: уровень урожая зависит от количества того минерального вещества, которое находится в минимуме. При этом увеличение содержания других минеральных веществ не обеспечивает увеличения урожая. Буссенго поставил ряд точных экспериментов, в результате которых доказал, что растения не используют азот, содержащийся в воздухе. Установив, что клевер и люцерна обогащают почву азотом, Буссенго предположил, что эти растения поглощают азот из воздуха. 2. Содержание минеральных элементов в растениях. Классификация минеральных элементов: макро- и микроэлементы. Зольные элементы сосредоточены в тех органах и клетках, уровень жизнедеятельности которых достаточно высок. Как правило, чем богаче почва и суше климат, тем выше содержание золы в растении. Необходимые элементы. Это те, без которых организм не может завершить свой жизненный цикл. Ю. Сакс и И. Кноп установили, что для жизнедеятельности растения, кроме С, О, Н, необходимы следующие 7 элементов: N, Р, S, К, Са, Mg, Fe. Позже была показана необходимость для растений в еще шести элементов: бора, марганца, цинка, меди, молибдена и хлора. Для некоторых растений характерны еще и другие элементы (например натрий). Все необходимые элементы корневого питания подчиняются основным правилам Либиха. Все необходимые для жизни растений элементы в зависимости от их количественного содержания в растении принято разделять на макроэлементы (содержание более 0,01%) — N, Р, S, К, Са, Mg, Fe и микроэлементы (содержание менее 0,01%) — Mn, Си, Zn, В, Mo, О Общие значения питательных элементов: 1) входят в состав биологически важных органических веществ; 2) участвуют в создании определенной ионной концентрации, стабилизации макромолекул и коллоидных частиц (электрохимическая роль); 3) участвуют в каталитических реакциях, входя в состав или активируя отдельные ферменты. 3. Макроэлементы – K, Ca, Mg, S, P, их физиологическая роль. Сера. Сера усваивается растениями только в окисленной форме—в виде аниона SCV. В растении основная масса аниона сульфата восстанавливается до —SH и —S—S— групп. Входит в состав трех аминокислот — цистина, цистеина и метионина, витаминов, коферментов (биотин, тиамин, коэнзим А, глютатион, липоевая кислота). Участвует в белковом обмене, аэробной фазе дыхания, синтезе жиров, в образовании макроэргической связи, в метаболизме и др. Часть серы находится а клеточном соке в виде иона сульфата. При отмирании корня в условиях, когда ему не хватает кислорода, серосодержащие соединения распадаются с образованием сероводорода, который ядовит для корня. При недостатке серы, так же как и при недостатке азота, начинается разрушение хлорофилла, но первыми испытывают недостаток серы верхние листья. Фосфор. Фосфор усваивается растениями в окисленной форме в виде солей фосфорной кислоты. Входит в состав нуклеиновых кислот (ДНК и РНК), нуклеотидов (АТФ, НАД, НАДФ), нуклеопротеидов, витаминов, фосфолипидов (мембраны) и др. Участвует в процессах гликолиза и аэробного дыхания. Освобождающаяся в этих процессах энергия накапливается в виде богатых энергией фосфатных связей. Принимает участие в фотосинтезе. При отсутствии фосфора а начале жизни и при последующей подкормке растения фосфорными солями листья растений некоторое время страдают из-за усиленного поступления фосфора и нарушенного в связи с этим азотного обмена. Калий, кальций и магний усваиваются из любых растворимый солей, анионы которых не обладают токсическим действием или связанные с каким-нибудь нерастворимым веществом, обладающим кислотными свойствами. Они (K, Ca, Mg) необходимы для питания. Роль: адсорбируясь на поверхности коллоидных частиц протоплазмы, они создают вокруг них определенные электростатические силы, они определяют оводненность и водоудерживающую силу коллоидов. Калий. Калий в растениях содержится в больших кол-ах, особенно в их вегетативных частях. Калий оказывает большое влияние на коллоиды плазмы, он повышает их гидрофильность (разжижает плазму). Калий является также катализатором ряда синтетических процессов: как правило, он катализирует синтез высокомолекулярных веществ из более простых, способствует синтезу сахарозы, крахмала, жиров, белков, При недостатке калия процессы синтеза нарушаются, и в растении скапливаются глюкоза, аминокислоты и продукты распада других высокомолекулярных соединений. При недостатке калия на нижних листьях появляется краевой запал — края листовой пластинки отмирают, листья приобретают характерную куполообразную форму, на листьях появляются некрозы. |