Генетика. кр номер 2. Закон чистоты гамет и его цитологическое обоснование. Второе правило Менделя. Особенности расщепления по фенотипу и генотипу. 3
 Скачать 1.62 Mb. Скачать 1.62 Mb.
|

8. Дать определение и привести примеры следующих явлений: множественные аллели, множественное действие гена, множественные гены.Множественные аллели – различное фенотипическое проявление аллелей одного и того же гена (характеризуют разнообразие генофонда вида). Обозначаются множественные аллели одной буквой с указанием номера: А, а1, а2, а3, а4 возникают они в результате многократных генных мутаций одного локуса гена. Чем больше аллельных генов, тем больше их комбинаций попарно. Например, наследование групп крови по антигенам АВ. I – ген определяющий развитие групп крови по системе АВО. Множественные аллели групп крови по системе АВО обозначаются: IO, IA , IB. А и В – агглютиногены – сложные соединения, за синтез которых отвечает ген «I». По наличию и сочетанию агглютиногенов в крови различают четыре группы крови по системе АВО. Плейотропия (плейотропность) — это множественное действие гена; тип наследования, при котором один ген определяет проявление нескольких признаков. Это свойство генов было хорошо исследовано на примере карликовости у мышей. Было показано, что при скрещивании фенотипически нормальных мышей в потомстве F1 мышки оказались карликовыми, из чего был сделан вывод, что карликовость обусловлена рецессивным геном. Рецессивные Действие генов-модификаторов гомозиготы прекращали расти на второй неделе, были неспособны к размножению, внутренние органы, особенно железы внутренней секреции, имели измененную форму, мыши были менее подвижны и плохо переносили перепады температур. Ген карликовости определял ненормальное развитие гипофиза, который, в свою очередь, определял раннюю остановку роста (изменение пропорций тела), ненормальное развитие половых желез (следовательно, стерильность), ненормальное развитие щитовидной железы, которое определяло пониженный обмен веществ, поэтому карликовые мыши были чувствительны к холоду, но более стойкие к голоду. Это цепочка последовательного изменения признаков при дефекте только одного гена. Множественные гены - неаллельные гены, контролирующие количественную (полигенную) генетическую изменчивость и взаимодействующие по типу некуммулятивной полимерии. При некумулятивной полимерии проявление признака не зависит от числа доминантных аллелей. Признак будет проявляться, если есть хотя бы один доминантный ген. При подобном эффекте наблюдается расщепление в потомстве в соотношении 15:1. 9. Наследование АВО системы групп крови 10.Наследование системы крови резус-факторР 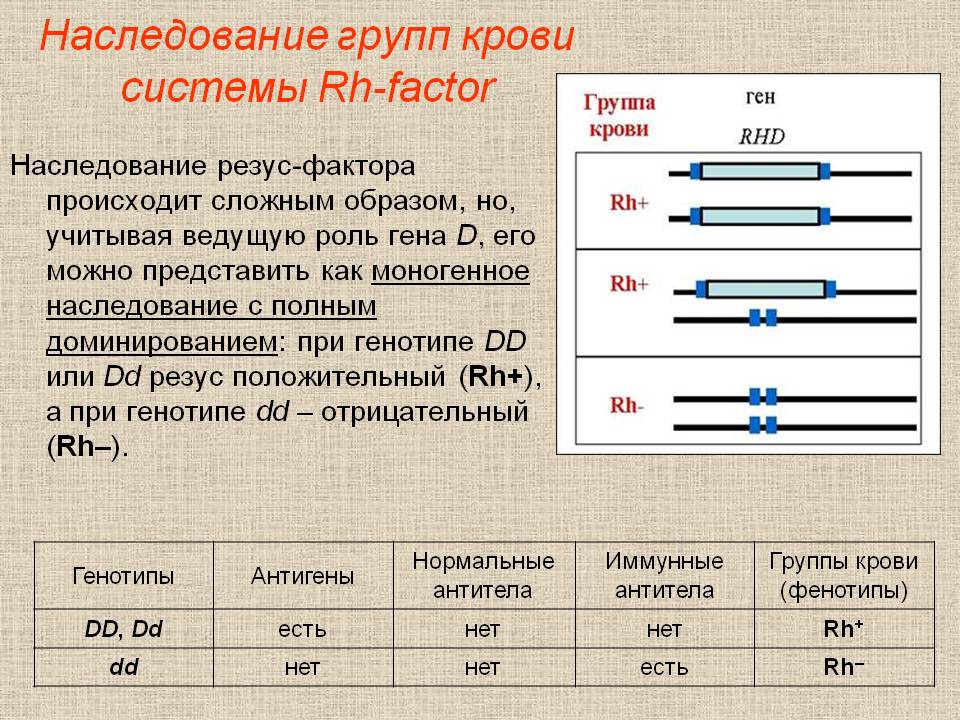 езус-фактор – Rh-фактор – антиген, содержащийся в эритроцитах человека. По наличию или отсутствию определяют резус положительный или резус отрицательный организм. езус-фактор – Rh-фактор – антиген, содержащийся в эритроцитах человека. По наличию или отсутствию определяют резус положительный или резус отрицательный организм. Резус – фактор передаётся по наследству и не изменяется в течение жизни Контролируется тремя парами аллельных генов Cc Dd Ee. Наибольшее значение имеет Dd РФ генетически детерминирован и наследуется по доминантному типу. Резус-положительный организм может иметь генотип DD или Dd, а отрицателный dd 11. Типы взаимодействия неаллельных генов. Неаллельные гены — это гены, расположенные в различных участках хромосом и кодирующие неодинаковые белки. Неаллельные гены также могут взаимодействовать между собой. Выделяют следующие типы взаимодействия неаллельных генов: комплементарность, эпистаз, полимерия, действие генов модификаторов, плейотропия. Комплиментарность: В том случае, когда признак появляется только при сочетании двух доминантных аллелей разных генов (например, А и В) их взаимодействие называют комплементарностью, а сами гены комплементарными (дополняющими друг друга). При этом каждый из взаимодействующих неаллельных генов в отсутствии другого не обеспечивает формирования признака. Известным примером комплементарного взаимодействия неаллельных генов является наследование окраски цветков душистого горошка при скрещивании двух родительских форм с белыми цветками ААвв и ааВВ. В потомстве F1 (АаВв), а также в F2 (фенотипический класс А-В-) будет появляться новая окраска – пурпурная. При этом в F2 соотношение классов с окрашенными цветками (А-В-) и классов с неокрашенными цветками (Аbb; ааВ- и ааbb) будет соответствовать формуле 9 : 7 Расщепление признаков при комплементарном взаимодействии генов 9 : 6 : 1 у тыквы , 9 : 3 : 4 Появление у льна , 9 : 3 : 3 : 1 Появление у дрозофилы потомства с темно-красными глазами ЭПИСТАЗ Эпистаз- взаимодействие двух неаллельных генов, при котором один ген, называемый эпистатичным или геном-супрессором, подавляет действие другого гена, называемого гипостатичным. Гены-супрессоры обозначаются I в случае доминантного состояния генов и i для их рецессивных аллелей. В настоящее время эпистаз разделяют на два типа: доминантный и рецессивный. Доминантный эпистаз Под доминантным эпистазом понимают подавление доминантным аллелем одного гена, действия аллельной пары другого гена. (А >В или В >А). Эпистатическая система обнаружена у кур. Некоторые породы кур имеют белое оперение (белый леггорн, виандотт и др.), другие же породы имеют окрашенное оперение. Белое оперение разных пород кур определяется несколькими различными генами. Так, например, доминантная белая окраска определяется генами CCII (белые леггорны), а рецессивная белая – ccii (белые виандотты). Ген С определяет наличие предшественника пигмента (хромогена), т. е. окрашенность пера, его аллель с – отсутствие хромогена и, следовательно, неокрашенность пера птицы. Ген I является подавителем действия гена С, аллель i не подавляет его действия. В присутствии даже одной дозы гена I в генотипе птицы действие генов окраски не проявится. Поэтому при скрещивании белых леггорнов (CCII) с белыми виндоттами (ccii) гибриды F1 оказываются белыми (СсIi). При скрещивании между собой гибридов F1 во втором поколении имеет место расщепление по окраске в отношении 13 белых: 3 окрашенных Рецессивный эпистаз. Под рецессивным эпистазом понимают такой тип взаимодействия, когда рецессивный аллель одного гена, будучи в гомозиготном состоянии, не дает возможности проявиться доминантному или рецессивному аллелям другого гена: аа>В; аа>bb или bb>А; bb>аа. Примером рецессивного эпистаза является окраска шерсти у собак породы лабрадор. Пигментация шерсти обеспечивается геном В, который в доминантном состоянии дает черную масть, а в рецессивном (b) – коричневую. Имеется также ген Е, который в доминантном состоянии не влияет на проявление окраски, но будучи в рецессивном состоянии (ее) подавляет синтез пигмента как черного, так и коричневого. Такие собаки становятся белыми. Расщепление в F2 будет следующее:  ПОЛИМЕРИЯ Полимери́я — взаимодействие неаллельных множественных генов, однонаправленно влияющих на развитие одного и того же признака; степень проявления признака зависит от количества генов. Такие свойства организмов, как рост, масса, количество молока и его жирность у скота, нельзя разложить на четкие фенотипические классы. Такие признаки необходимо оценивать в количественном выражении, поэтому они чаще всего называются количественными, или мерными. Пример: цвет кожи у человека определяют четыре пары неаллельных генов, ответственных за выработку пигмента меланина. Чем больше имеется доминантных генов, тем больше в коже меланина, и тем темнее её окраска. У человека с доминантными генами A1A1A2A2A3A3A4A4 кожа самая тёмная. У человека, в генотипе которогосодержатся только рецессивные гены a1a1a2a2a3a3a4a5, кожа самая светлая. В зависимости от соотношения числа доминантных и рецессивных генов цвет кожи изменяется от светлого до тёмного. Такой тип взаимодействия называется кумулятивной полимерией. При кумуляции степень проявления признака зависит от числа доминантных генов (чем больше генов, тем сильнее выражен признак). Потомство при кумулятивной полимерии разделяется следующим образом — 1:4:6:4:1 (у одной особи признак выражен максимально, у остальных наблюдается его угасание до полного исчезновения). При некумулятивной полимерии проявление признака не зависит от числа доминантных аллелей. Признак будет проявляться, если есть хотя бы один доминантный ген. При подобном эффекте наблюдается расщепление в потомстве в соотношении 15:1. ПЛЕЙОТРОПИЯ При плейотропном действии гена один ген определяет развитие или влияет на проявление нескольких признаков. Это свойство генов было хорошо исследовано на примере карликовости у мышей. Было показано, что при скрещивании фенотипически нормальных мышей в потомстве F1 мышки оказались карликовыми, из чего был сделан вывод, что карликовость обусловлена рецессивным геном. Рецессивные Действие генов-модификаторов гомозиготы прекращали расти на второй неделе, были неспособны к размножению, внутренние органы, особенно железы внутренней секреции, имели измененную форму, мыши были менее подвижны и плохо переносили перепады температур. Ген карликовости определял ненормальное развитие гипофиза, который, в свою очередь, определял раннюю остановку роста (изменение пропорций тела), ненормальное развитие половых желез (следовательно, стерильность), ненормальное развитие щитовидной железы, которое определяло пониженный обмен веществ, поэтому карликовые мыши были чувствительны к холоду, но более стойкие к голоду. Это цепочка последовательного изменения признаков при дефекте только одного гена. Алле́ли — различные формы одного и того же гена, расположенные в одинаковых участках гомологичных хромосом, определяют направление развития конкретного признака. ДЕЙСТВИЕ ГЕНОВ-МОДИФИКАТОРОВ Гены-модификаторы – это гены, влияющие на проявление признаков (количественных или качественных), контролируемых другими неаллельными генами. Примером действия генов-модификаторов является контроль за расположением окрашенных участков шерсти (пятен) на теле некоторых животных. При наличии доминантного гена S (генотип SS или Ss) окраска шерсти у мышей, морских свинок, собак, лошадей и других животных равномерная и пятнистость отсутствует. Если ген находится в рецессивном состоянии (генотип ss) – пятна интенсивно проявляются |