биохимия. Значение изучаемой темы
 Скачать 1.04 Mb. Скачать 1.04 Mb.
|

Тема № 3. Биологическое окисление. Определение активности каталазы Значение изучаемой темы: Тканевое дыхание обеспечивает энергией весь организм. Нарушение процессов тканевого дыхания приводит к гипоэнергетическим состояниям, которые являются причиной многих патологических состояний. а. 1. Основные понятия и положения темы1.1. Понятие о катаболизме, анаболизме, биологическом окисленииОкисление - это отщепление от вещества электронов; восстановление - это присоединение электронов. Метаболизм - это обмен веществ. Он имеет две стороны: катаболизм и анаболизм. Катаболизм - это расщепление сложных органических веществ до более простых с выделением энергии. Анаболизм - это синтез сложных веществ из простых веществ с использованием энергии. Для поддержания гомеостаза (постоянства внутренней среды) в организме постоянно вырабатывается энергия. Источником энергии является расщепление пищевых веществ (белков, жиров и углеводов), а также аналогичных молекул самого организма до СО2, Н2О и NН3 (для азотсодержащих веществ). Под биологическим окислением понимают все окислительно-восстановительные реакции, происходящие в организме. Наиболее важным видом окисления с энергетической точки зрения является дегидрирование субстратов с последующим взаимодействием отщепленного водорода с кислородом, полученным из атмосферы при дыхании, с образованием воды. В отличие от неживой природы, где энергия при горении веществ выделяется в виде света и тепла, в организме человека и животных энергия окислительно-восстановительных реакций используется на синтез АТФ, то есть переводится в химическую форму, а также выделяется в виде тепла. 1.2. Этапы катаболизма1-й этап. На этом этапе макромолекулы расщепляются до своих мономеров (или строительных блоков). Так, полисахариды распадаются до моносахаридов (гексоз и пентоз); жиры - до глицерина и жирных кислот; белки - до аминокислот. Этот этап является специфическим, так как каждая макромолекула (полимер) своим набором ферментов расщепляется до мономеров. 1-й этап катализируется ферментами класса гидролаз. Он локализован в пищеварительном тракте для пищевых (экзогенных) макромолекул, а для эндогенных (находящихся в клетках организма) - в основном в лизосомах. Этот этап энергетической ценности не имеет. В нем выделяется менее 1% энергии, которая рассеивается в виде тепла. 2-й этап. Является специфическим путем катаболизма. На этом этапе каждый из мономеров своим собственным путем превращается в одну из карбоновых кислот. Моносахариды, глицерин и некоторые аминокислоты превращаются в пируват. Жирные кислоты и часть аминокислот - в ацетил-КоА (активную уксусную кислоту). Некоторые аминокислоты превращаются в оксалоацетат или 2-оксоглутарат. 2-й этап происходит в митохондриях и цитозоле клеток. Образующаяся при этом энергия выделяется в виде тепла и используется на синтез АТФ. 3-этап. Является общим для разных классов веществ. На этом этапе пируват в процессе окислительного декарбоксилирования превращается в ацетил-КоА. Ацетил-КоА, оксалоацетат и 2-оксоглутарат окисляются в цикле Кребса. За один оборот цикла Кребса образуются 2 молекулы СО2,. Водород, полученный в дегидрогеназных реакциях, присоединяется к НАД+ и ФАД с образованием НАДН и ФАДН2, которые окисляются в дыхательной цепи. Приэтом образуется вода, а выделившаяся энергия используется на синтез АТФ и рассеивается в виде тепла. Все реакции этого этапа локализованы в митохондриях. 1.3. Окислительное декарбоксилирование пируватаОкислительное декарбоксилирование пировиноградной кислоты (пирувата) катализируется пируватдегидрогеназой (ПДГ). ПДГ - это комплекс из трех ферментов, для работы которого требуются следующие витамины: В1 (тиамин), В2 (рибофлавин), РР (никотинамид), пантотеновая и липоевая кислоты. ПДГ осуществляет декарбоксилирование (отщепление карбоксильной группы) и окисление (отщепление водорода) молекулы пирувата. СН3 СН3  С  =О + НS-КоА + НАД+ С=О + СО2 + НАДН + Н+ =О + НS-КоА + НАД+ С=О + СО2 + НАДН + Н+ СООН S-КоА Пируват Ацетил-КоА Продукты реакции и их дальнейшие превращения:СО2. В основном выделяется с выдыхаемым воздухом, а также используется для карбоксилирования субстратов. Ацетил-КоА. Расщепляется в цикле Кребса до СО2 и Н2; НАДН. Окисляется в дыхательной цепи. Регуляция ПДГ: ПДГ может быть в двух формах: фосфорилированной (неактивной) и дефосфорилированной (активной). АТФ, НАДН, ацетил-КоА, жирные кислоты ингибируют ПДГ, инсулин активирует. 1.4. Цикл Кребса (цикл трикарбоновых кислот, цикл лимонной кислоты)Ацетил-КоА, образованный в результате окисления жирных кислот и пировиноградной кислоты, в цикле Кребса распадается СО2 и Н2. Итоговое уравнение цикла Кребса: А 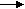 цетил-КоА + 3НАД+ + ФАД + 2Н2О + АДФ + Н3РО4 2СО2 + 3НАДН + ФАДН2 + АТФ цетил-КоА + 3НАД+ + ФАД + 2Н2О + АДФ + Н3РО4 2СО2 + 3НАДН + ФАДН2 + АТФВ ходе цикла Кребса образуется сукцинил-КоА, который содержит макроэргическую связь и является первичным макроэргом. Гидролиз макроэргической связи сопряжен с реакцией фосфорилирования АДФ, при этом образуется АТФ. Синтез АТФ за счет энергии, накопленной в субстрате, называется субстратным фосфорилированием.  Конечные продукты цикла Кребса и пути их использования:- СО2 выдыхается с воздухом, небольшая часть используется в реакциях карбоксилирования. - НАДН и ФАДН2 окисляются в дыхательной цепи. - АТФ используется на различные виды работы: 1) механическая работа (сокращение мышц, движение сперматозоидов, лейкоцитов); 2) осмотическая работа или активный транспорт, то есть движение против градиента концентрации; 3) химическая работа, когда энергия АТФ используется в биосинтетических процессах и на активацию субстратов; 4) электрическая работа (генерация биотоков); 5) при передаче гормонального сигнала (для работы аденилатциклазы и протеинкиназы). Регуляция цикла КребсаОсуществляется путем влияния на ключевые ферменты: цитратсинтазу (начинает процесс), изоцитратдегидрогеназу (лимитирующий фермент), 2-оксоглутаратдегидрогеназу (фермент, стоящий на развилке). Цитратсинтазу активируют оксалоацетат и ацетил-КоА, ингибируют АТФ, НАДН, длинноцепочные ацилы-КоА, сукцинил-КоА. Изоцитратдегидрогеназа является аллостерическим ферментом. Ее активируют АДФ, Са2+, цАМФ. Ингибируют изоцитратдегидрогеназу АТФ, НАДН, НАДФН. 2-оксоглутаратдегидрогеназу активируют Са2+ и цАМФ, ингибирует сукцинил-КоА. Цикл Кребса активируется под влиянием катехоламинов, глюкагона и йодтиронинов. Значение цикла Кребса:- катаболическое и энергетическое (цикл Кребса является общим конечным путем распада для метаболитов всех классов соединений; в нем образуется АТФ в результате субстратного фосфорилирования; он является главным поставщиком водорода для дыхательной цепи); - анаболическое или биосинтетическое. Промежуточные метаболиты цикла Кребса используются на синтез других соединений. Например, из оксалоацетата, 2-оксоглутарата и сукцината образуются аминокислоты; из оксалоацетата - глюкоза и другие углеводы; сукцинил-КоА используется на синтез гема; - регуляторное. Метаболиты - цитрат и АТФ являются регуляторами других процессов. Они активируют синтез жирных кислот и ингибируют гликолиз. 1.5. Дыхательная цепьДыхательная цепь (цепь переноса электронов) - это цепь сопряженных окислительно-восстановительных реакций, в ходе которых водород, отщепленный от субстратов, переносится на кислород с образованием воды и выделением энергии. Назначение дыхательной цепи - генерирование энергии. Компоненты дыхательной цепи называются дыхательными переносчиками. Большинство из них (кроме убихинона) являются сложными белками. Схема дыхательной цепиСубстраты НАД-зав. Субстраты ФАД-зав. дегидрогеназ дегидрогеназ 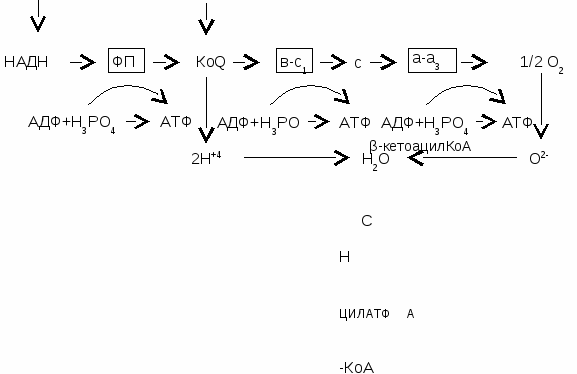 Характеристика дыхательных переносчиков:НАДН-дегидрогеназа (НАДН-ДГ) (в схеме - ФП) - это флавинзависимый фермент, небелковой частью которого является ФМН и железо-серные центры. НАДН-ДГ встроена во внутреннюю мембрану митохондрий. Она осуществляет перенос водорода с НАДН вначале на ФМН с образованием ФМНН2, затем переносит водород с ФМНН2 на железо-серные центры и только потом на КоQ, при этом последний восстанавливается до КоQН2. Таким образом, НАДН-ДГ катализирует реакцию: НАДН-ДГ  НАДН2 + КоQ НАД+ + КоQН2 НАДН2 + КоQ НАД+ + КоQН2 КоQ (убихинон) - это небелковый переносчик, растворимый в липидах. Восстановленная форма убихинона (КоQН2) называется убихинол. Убихинон может перемещаться в липидной фазе внутренней мембраны митохондрий, представляя, таким образом, лабильный субстрат для ферментов встроенных в мембрану. Цитохромы (b, c1, c, a, а3) - это сложные белки, небелковой частью которых является гем, содержащий Fe3+. Принимая электрон, железо трехвалентное переходит в железо двухвалентное, отдавая электрон - переходит опять в трехвалентное.  Fe3+ + e- Fe2+ Fe3+ + e- Fe2+ Fe2+ - e- Fe3+ Fe2+ - e- Fe3+Комплекс цитохромов b-c1 является ферментом (КоQН2 -дегидро-геназой). Он переносит электроны с КоQН2 на цитохром c, при этом железо цитохрома восстанавливается до двухвалентного. Протоны атомов водорода выбрасываются в межмембранное пространство. Таким образом, осуществляется реакция: КоQН2  КоQН2 + 2c(Fe3+) KoQ +2Н+ + 2c(Fe2+) КоQН2 + 2c(Fe3+) KoQ +2Н+ + 2c(Fe2+)Комплекс цитохромов a-a3 является оксидазой. Он переносит электроны с цитохрома c на кислород, превращая последний в ион (О2-). Цитохромоксидаза катализирует реакцию: а-а3 aa3  2c(Fe2+) + 1/2 О2 2c(Fe3+) + О2- 2c(Fe2+) + 1/2 О2 2c(Fe3+) + О2- Ионы кислорода и протоны водорода взаимодействуют с образованием воды. Перенос электронов по дыхательной цепи происходит по градиенту окислительно-восстановительного потенциала (Ео). Окислительно-восстановительный потенциал характеризует способность сопряженной окислительно-восстановительной пары обратимо отдавать электроны. Чем более отрицательна величина Ео, тем выше способность данной пары отдавать электроны, чем более положительна - тем выше способность принимать электроны. Величина Ео у пары НАДН/НАД+ -0,32в, Ео у пары Н2О/О2- +0,82в, при этом разность между этими величинами составляет 0,82-(-0,32)=1,14в. Этому соответствует разность свободной энергии - 220 кдж на пару переносимых электронов. Этого количества энергии достаточно для синтеза 4 молекул АТФ. Однако в дыхательной цепи синтезируется только 3 АТФ. Синтез АТФ происходит в тех участках дыхательной цепи, где имеет место наибольший перепад окислительно-восстановительного потенциала. В этих участках энергии выделяется столько, что ее достаточно для проведения реакции фосфорилирования АДФ.  АДФ + Н3РО4 АТФ АДФ + Н3РО4 АТФТаким образом, в дыхательной цепи синтез АТФ (фосфорилирование) энергетически сопряжен с переносом электронов, то есть окислением дыхательных переносчиков. Синтез АТФ, сопряженный с переносом электронов по дыхательной цепи, называется окислительным фосфорилированием. Участки дыхательной цепи, где есть такой синтез, называют пунктами сопряжения окисления с фосфорилированием. Пункты сопряжения: между НАДН и КоQ, на участке цитохромов b - c1, и a - a3. Таким образом, окисление 1 молекулы НАДН приводит к синтезу 3 молекул АТФ, окисление 1 молекулы ФАДН2 - к образованию 2 молекул АТФ. Для работы цикла Кребса и дыхательной цепи требуются следующие витамины: В1, В2, РР, Q, пантотеновая и липоевая кислоты. 1.6. Механизм окислительного фосфорилированияКомпоненты электрон-транспортной цепи (дыхательной цепи) находятся во внутренней мембране митохондрий (непроницаемой для протонов водорода). Они расположены таким образом, что, передавая электроны по цепи, одновременно выталкивают протоны водорода на наружную сторону мембраны в межмембранное пространство. В результате, на наружной стороне мембраны создается избыток протонов водорода (положительный заряд), а с внутренней стороны – недостаток (отрицательный заряд). Это проявляется в возникновении мембранного электрохимического потенциала Н+, который складывается из разности зарядов на мембране () и разности рН (снаружи более кислое, внутри – более щелочное). Протоны водорода могут возвращаться в матрикс по протонным каналам, с которыми связана специфическая Н+-АТФаза (АТФ-синтаза). Обратный ток протонов по каналам активирует этот фермент, и он катализирует синтез АТФ из АДФ и Н3РО4. Таким образом, энергия мембранного поценциала преобразуется в энергию макроэргической связи АТФ. АТФ с помощью фермента транслоказы переносится из митохондрий в цитозоль, где и используется. 1.7. Сопряжение и разобщение в дыхательной цепиСопряжение в дыхательной цепи - это такое состояние, когда окисление (перенос электронов) сопровождается фосфорилированием, то есть синтезом АТФ. Разобщение - это такое состояние дыхательной цепи, когда окисление идет, а фосфорилирование не происходит, то есть пункты фосфорилирования выключены полностью или частично. В этом случае вся или какая-то часть образующейся энергии выделяется в виде тепла. Сопряженность дыхательной цепи можно оценить по коэффициенту Р/О. Коэффициент Р/О равен числу молей АТФ, образующихся из АДФ и Н3РО4 , на 1 грамм-атом поглощенного кислорода. Разобщение в дыхательной цепи могут вызывать липофильные вещества, которые способны переносить протоны водорода с внешней стороны внутренней мембраны митохондрий на внутреннюю, минуя АТФ-синтазу. В результате вся энергия электрохимического потенциала будет рассеиваться в виде тепла. Разобщение вызывают: 2,4-ДНФ (динитрофенол), многие яды промышленных производств, бактериальные токсины, набухание митохондрий, жирные кислоты, ионофоры (вещества, переносящие ионы через мембрану). Разобщители повышают скорость переноса электронов по дыхательной цепи и выводят ее из под ингибирующего влияния АТФ. Регуляция дыхательной цепи1. АДФ стимулирует работу дыхательной цепи. Это явление называется дыхательным контролем. 2. АТФ тормозит работу дыхательной цепи и потребление кислорода. 3. Адреналин и глюкагон активируют работу дыхательной цепи. Блокаторы дыхательной цепи1. Ротенон блокирует дыхательную цепь на участке НАДН – КоQ. 2. Амитал, антимицин - на участке между цитохромами b и c1. 3. Цианиды и окись углерода блокируют цитохромоксидазу, при этом вся дыхательная цепь не работает. 1.8. Нефосфорилирующее (свободное) окислениеЭто окисление без образования АТФ. Ферменты свободного окисления: оксидазы, оксигеназы, некоторые дегидрогеназы. Значение свободного окисления: - терморегуляция; - образование биологически важных соединений (катехоламинов, глюкокортикостероидов, коллагена, активация витамина Д и т.д); - обезвреживание ксенобиотиков (ядов, токсинов, лекарств, веществ бытовой химии). 1.9. Тканевые и возрастные особенности окислительных процессовАнаэробные ткани могут получать энергию без кислорода. К анаэробным тканям относятся скелетные мышцы, эритроциты, периферические нервы, мозговое вещество почек, кость, хрящ, соединительная ткань. Аэробные ткани получают энергию с использованием кислорода и полностью зависят от кровотока. Аэробными тканями являются головной мозг, сетчатка глаза, сердце, кора почек, печень, слизистая тонкого кишечника. Потребление кислорода, а значит, и интенсивность окислительных процессов с возрастом падают. Диагностическое значение определения активности каталазы Каталаза содержится во всех тканях и жидкостях организма, но особенно много ее в строме эритроцитов и печени. В процессе окисления некоторых веществ образуется пероксид водорода, ядовитый для организма. Каталаза расщепляет пероксид водорода на молекулярный кислород и воду. Определение активности каталазы имеет значение для диагностики рака, анемии, туберкулеза. При этих заболеваниях активность каталазы снижается. б) Ситуационные задачи 1. К препарату митохондрий добавили пируват, меченый 14С по метильной группе. Какое положение займет 14С в оксалоацетате после одного оборота цикла Кребса? 2. Сколько молекул АТФ синтезируется при окислении одной молекулы пирувата до 2-оксоглутарата? Одной молекулы изоцитрата до сукцината? Одной молекулы сукцината до оксалоацетата? При условии, что дегидрогеназные реакции сопряжены с дыхательной цепью. 3. Ротенон (токсичное вещество, вырабатываемое одним из видов растений) резко подавляет активность митохондриальной НАДН-дегидрогеназы. Токсичный антибиотик антимицин сильно ингибирует окисление убихинола. Допустим, что оба эти вещества блокируют соответствующие участки дыхательной цепи с равной эффективностью. Какой из них будет при этом более мощным ядом? Дайте аргументированный ответ. 4. Напишите реакции, которые ускоряются при введении в организм кокар-боксилазы (активной формы витамина В1). 5. Подберите каждому пронумерованному ферменту дыхательной цепи соот-ветствующий кофермент, обозначенный буквами: 1. НАДН-дегидрогеназа А. ФАД 2. QН2 - дегидрогенза Б. гем 3. Цитохромоксидаза В. ФМН 4. Малатдегидрогеназа Г. гем, Сu2+ 5. Сукцинатдегидрогеназа Д. НАД+ Е. НАДФ+ 6. Как отразится передозировка барбитуратами на потребление кислорода? 7. 2,4-ДНФ (сильный разобщающий агент) пытались одно время исполь-зовать для борьбы с ожирением. Теперь такого рода разобщающие агенты уже не применяются в качестве лекарственных препаратов, поскольку известны случаи, когда их применение приводило к летальному исходу. Почему прием разобщающих агентов может вызвать смерть? 8. Какую физиологическую функцию определяет величина Р/О? |