физиология экзамен. Физиология мои ответы экзамен. 40. Детектирование сигналов и опознание образов. Детектирование сигналов
 Скачать 4.25 Mb. Скачать 4.25 Mb.
|

|
40. Детектирование сигналов и опознание образов. Детектирование сигналов - это избирательное выделение сенсорным нейроном того или иного признака раздражителя, имеющего поведенческое значение. Такой анализ осуществляют нейроны-детекторы, избирательно реагирующие лишь на определенные параметры стимула. Так, типичный нейрон зрительной области коры отвечает разрядом лишь на одну определенную ориентацию темной или светлой полоски, расположенной в определенной части поля зрения. При других наклонах той же полоски ответят другие нейроны. В высших отделах сенсорной системы сконцентрированы детекторы сложных признаков и целых образов. Примером могут служить детекторы лиц в нижневисочной области коры мозга. Опознание образов - конечная и наиболее сложная операция сенсорной системы. Она заключается в отнесении образа к тому или иному классу объектов, с которыми ранее встречался организм, т. е. в классификации образов. Синтезируя, т.е. объединяя сигналы от нейронов-детекторов, высший отдел сенсорной системы формирует «образ» раздражителя и сравнивает его с множеством образов, хранящихся в памяти. Опознание завершается принятием решения о том, с каким объектом или ситуацией встретился организм. В результате этого происходит восприятие, т. е. мы осознаем, чье лицо видим перед собой, кого слышим, какой запах чувствуем. Опознание часто происходит независимо от изменчивости сигнала. Мы надежно опознаем, например, предметы при различной их освещенности, окраске, размере, ракурсе, ориентации положении в поле зрения. Это означает, что сенсорная система формирует независимый от изменений ряда признаков сигнала (инвариантный) сенсорный образ. 41. Механизмы переработки информации в сенсорных системах. Переработка информации в сенсорной системе осуществляется процессами возбудительного и тормозного взаимодействия нейронов. Возбудительное взаимодействие заключается в том, что аксон каждого нейрона, приходя в вышележащий слой сенсорной системы, активирует несколько нейронов, каждый из которых в свою очередь получает сигналы от нескольких клеток предыдущего слоя. Совокупность рецепторов, сигналы которых поступают на данный нейрон, называют его рецептивным полем. Рецептивные поля соседних нейронов частично перекрываются. В результате такой организации связей в сенсорной системе образуется так называемая нервная сеть. Благодаря ей повышается чувствительность системы к слабым сигналам, а также обеспечивается высокая приспособляемость к меняющимся условиям среды. Тормозная переработка сенсорной информации основана на том, что каждый возбужденный сенсорный нейрон активирует тормозной интернейрон. Интернейрон в свою очередь подавляет импульсацию как самого возбудившего его элемента (последовательное, или возвратное, торможение), так и его соседей по слою (боковое, или латеральное, торможение). Сила этого торможения тем больше, чем сильнее возбужден первый элемент и чем ближе к нему соседняя клетка. Значительная часть операций по снижению избыточности и выделению наиболее существенных сведений о раздражителе производится латеральным торможением. 42.Адаптация сенсорных систем. Сенсорная система обладает способностью приспосабливать свои свойства к условиям среды и потребностям организма. Сенсорная адаптация - общее свойство сенсорных систем, которое заключается в приспособлении к длительно действующему (фоновому) раздражителю. Адаптация проявляется в снижении абсолютной и повышении дифференциальной чувствительности сенсорной системы. Субъективно адаптация проявляется в привыкании к действию постоянного раздражителя (например, мы не замечаем непрерывного давления на кожу привычной одежды). Адаптационные процессы начинаются на уровне рецепторов, охватывая и все нейронные уровни сенсорной системы. Адаптация выражена слабо только в вестибуло- и проприорецепторах. По скорости данного процесса все рецепторы делятся на быстро- и медленно адаптирующиеся. Первые после развития адаптации практически не посылают в мозг информации о длящемся раздражении. Вторые эту информацию передают в значительно ослабленном виде. Когда действие постоянного раздражителя прекращается, абсолютная чувствительность сенсорной системы восстанавливается. Так, после выключения света абсолютная чувствительность зрения в темноте резко повышается. В сенсорной адаптации важную роль играет эфферентная регуляция свойств сенсорной системы. Она осуществляется за счет нисходящих влияний более высоких на более низкие ее отделы. Происходит как бы перенастройка свойств нейронов на оптимальное восприятие внешних сигналов в изменившихся условиях. Состояние разных уровней сенсорной системы контролируется также ретикулярной формацией, включающей их в единую систему, интегрированную с другими отделами мозга и организма в целом. Эфферентные влияния в сенсорных системах чаще всего имеют тормозной характер, т.е. приводят к уменьшению их чувствительности и ограничивают поток афферентных сигналов. 43.Взаимодействие сенсорных систем. ( маловато, но норм инфы больше нема) Взаимодействие сенсорных систем осуществляется на спинальном, ретикулярном, таламическом и корковом уровнях. Особенно широка интеграция сигналов в ретикулярной формации. В коре большого мозга происходит интеграция сигналов высшего порядка. В результате образования множественных связей с другими сенсорными и неспецифическими системами многие корковые нейроны приобретают способность отвечать на сложные комбинации сигналов разной модальности. Это особенно свойственно нервным клеткам ассоциативных областей коры большого мозга, которые обладают высокой пластичностью, что обеспечивает перестройку их свойств в процессе непрерывного обучения опознанию новых раздражителей. Межсенсорное (кросс-модальное) взаимодействие на корковом уровне создает условия для формирования «схемы мира» и непрерывной увязки, координации с ней собственной «схемы тела» организма. 44.Обонятельная сенсорная система, ее функции: - Периферический отдел обонятельной сенсорной системы; Периферическим отделом обонятельной сенсорной системы являются рецепторные клетки в эпителиальной (обонятельной) выстилке носовой полости. Она расположена в верхней носовой раковине и соответствующей части носовой перегородки, отличается желтоватым цветом (из-за присутствия пигмента в клетках) и занимает в носовой полости около 2,5–5 см2. Слизистая оболочка носовой полости в области обонятельной выстилки несколько утолщена по сравнению с остальной слизистой. Она образована рецепторными и опорными клетками. Обонятельные рецепторные клетки представляют собой первичночувствующие клетки. В их апикальной части расположен длинный тонкий дендрит, заканчивающийся булавовидным утолщением. От утолщения отходят многочисленные реснички, имеющие обычное строение и погруженные в слизь. Эту слизь выделяют опорные клетки и железы, лежащие под эпителиальным пластом (Боуменовы железы). В базальной части клетки расположен длинный аксон. Немиелинизированные аксоны многих рецепторных клеток образуют под эпителием довольно толстые пучки, называемые обонятельными волокнами (fila olfactoria). Эти аксоны проходят в отверстия продырявленной пластинки решетчатой кости и направляются к обонятельной луковице, лежащей на нижней поверхности головного мозга. Возбуждение рецепторных клеток происходит при взаимодействии стимула с ресничками, затем оно по аксону передается в мозг. Хотя обонятельные клетки являются нейронами, они, в отличие от последних, способны к обновлению. Срок жизни этих клеток составляет примерно 60 дней, после чего они дегенерируют и фагоцитируются. Замена рецепторных клеток происходит за счет деления базальных клеток обонятельной выстилки. - Механизм возбуждения обонятельных рецепторов; Присоединение пахучих молекул к хеморецептивным белкам запускает каскад биохимических реакций, приводящих к возникновению потенциалов действия рецепторных клеток. После присоединения пахучих молекул к рецепторам происходит активация G-белков, а затем аденилатциклазы, что ведет к повышению внутриклеточной концентрации циклического аденозинмонофосфата, который активирует протеинкиназу А. Ее фосфорилирующее действие на белки мембранных каналов для катионов приводит к открытию каналов, току катионов в клетку и деполяризации мембраны, являющейся рецепторным потенциалом первичного сенсорного нейрона. При деполяризации мембраны до критического уровня в сенсорном нейроне возникают потенциалы действия, служащие для передачи информации на следующий иерархический уровень обонятельной системы. Все первичные обонятельные нейроны имеют широкий спектр чувствительности и реагируют на многие пахучие вещества, в то же время разные рецепторные клетки отличаются индивидуальной чувствительностью к различным веществам (обонятельный профиль сенсорного нейрона). Каждый запах возбуждает одновременно большое количество рецепторов, индивидуальные особенности которых в совокупности создают неповторимую комбинацию, характерную именно для этого запаха. На этом основана способность человека к различению сложных запахов, принадлежащих, например, двум разновидностям духов. Проводниковый_и_корковый_отделы_обонятельной_сенсорной_системы.'>- Проводниковый и корковый отделы обонятельной сенсорной системы. Проводниковый отдел представлен проводящими нервными путями в виде обонятельного нерва, ведущие к обонятельной луковице (образование овальной формы). Проводниковыйотдел. Первым нейроном обонятельного анализатора следует считать нейросенсорную или нейрорецепторную клетку. Аксон этой клетки образует синапсы, называемые гломерулами, с главным дендритом митральных клеток обонятельной луковицы, которые представляют второй нейрон. Аксоны митральных клеток обонятельных луковиц образуют обонятельный тракт, который имеет треугольное расширение (обонятельный треугольник) и состоит из нескольких пучков. Волокна обонятельного тракта отдельными пучками идут в передние ядра зрительного бугра. Центральный отдел состоит из обонятельной луковицы, связанной ветвями обонятельного тракта с центрами, которые расположены в палеокортексе (древней коре больших полушарий головного мозга) и в подкорковых ядрах, а также корковый отдел, который локализован в височных долях мозга, извилине морского коня. Центральный, или корковый, отдел обонятельного анализатора локализуется в передней части грушевидной доли коры в области извилины морского коня. Безмякотные аксоны биполярных рецепторных клеток объединяются в 15—20 пучков, каждый из которых содержит несколько сотен или тысяч волокон. Такие пучки входят в полость черепа через отверстия решетчатой кости и объединяются в обонятельные нервы (I пара черепных нервов). Окончания первичных сенсорных нейронов образуют синапсы с дендритами митральных клеток обонятельных луковиц. Обонятельные луковицы расположены в передней черепной ямке над решетчатой костью, их наружный слой образован сплетениями волокон обонятельных нервов. Чуть глубже, в клубочковом слое, информация передается митральным клеткам, причем каждая митральная клетка (сенсорный нейрон второго порядка) получает в одном клубочке сигналы приблизительно тысячи аксонов рецепторных клеток. Митральным клеткам свойственна ритмическая активность, изменения которой обусловлены вдыханием пахучих веществ. Локальные интернейроны обонятельных луковиц (перигломерулярные и зернистые клетки) осуществляют латеральное торможение, способствующее контрастированию передаваемых сигналов. На локальных интернейронах оканчиваются эфферентные волокна противоположной обонятельной луковицы, лимбических структур и ретикулярной формации ствола: эфферентное влияние позволяет контролировать характер выходных сигналов обонятельной луковицы. Аксоны митральных клеток вместе с прилегающими к ним аксонами пучковых клеток, которые получают афферентную информацию от нескольких клубочков одновременно, образуют латеральный обонятельный тракт к центрам высшего порядка. Латеральный обонятельный тракт разделяется на несколько частей, оканчивающихся в лимбических структурах переднего мозга: переднем обонятельном ядре, перегородке, пириформной и парагиппокампальной извилинах. Нейроны этих структур возбуждаются при поступлении афферентной информации от обонятельных рецепторов и передают ее гиппокампу, миндалинам, орбитофронтальной коре и ретикулярной формации среднего мозга. Большинство областей проекции обонятельного тракта не принимают непосредственного участия в восприятии запахов, их физиологическая роль состоит в образовании ассоциативных связей обонятельной системы с другими сенсорными системами при формировании пищевого, полового и оборонительного поведения. Активация лимбической системы создает эмоциональный компонент обонятельного восприятия, который определяет субъективное отношение к запахам. 46.Соматосенсорная система, ее функции (виды рецепторов кожи, их функции; механизм возбуждения механорецепторов кожи; механизм возбуждения терморецепторов кожи; интрафузальные мышечные веретена, их функция и механизм возбуждения; сухожильные рецепторы Гольджи, их функция и механизм возбуждения;. проводниковый отдел соматосенсорной системы; лемнисковый путь проведения соматосенсорной информации; спиноталамический путь проведения соматосенсорной информации; корковый отдел соматосенсорной системы, сенсорный гомункулюс, его функции.) В соматосенсорную анализаторную систему включают систему кожной чувствительности и чувствительную систему скелетно-мышечного аппарата, главная роль в которой принадлежит проприорецепции. Кожная рецепция (виды рецепторов кожи, виды кожной чувствительности). Виды рецепторов кожи: - Пластинчатые тельца. (тельца Фатера-Пачини) располагаются в соединительной ткани внутренних органов и глубоких слоях кожи. Соединительнотканные пластинки и интерстициальная жидкость капсулы способствуют усилению давления на нервное окончание, в результате чего аксолемма деформируется, изменяется её проницаемость и происходит генерация потенциала. Местом возникновения потенциала действия в тельцах Пачини является область первого перехвата Ранвье. - Осязательные тельца (Мейснера) размещаются в сосочковом слое дермы. Это тактильные рецепторы, реагирующие на прикосновение. Незначительная деформация капсулы передается глиоцитам и далее на дендриты. - Клетки Меркеля расположены под эпидермисом. Реагируют на внезапные смещения кожи, например, при поглаживании. - Колбы Краузе – локализованы в дерме и подкожной клетчатке у основания сухожилий. Осуществляют рецепцию локальных изменений давления и растяжения. - Механочувствительные свободные окончания в коже. Являются немиелинизированными аксонами, у них нет корпускулярных структур. К ним относятся рецепторы волосяных фолликул, реагирующих на давление от движения волоса. - Генитальные тельца Догеля – находятся в наружных половых органах, молочных железах. Реагируют на давление импульсами, вызывающими половое возбуждение. Виды кожной чувствительности: 1) болиевые 2) тепловые 3) холодовые 4) прикосновения и давления Согласно этой классификации, в основе разного характера кожных ощущений лежат различия в пространственном и временном распределении импульсов в афферентных волокнах, возбуждаемых при разных видах кожных раздражений. Исследования электрической активности одиночных нервных окончаний и волокон свидетельствуют о том, что многие из них воспринимают лишь механические или температурные стимулы. Тактильная рецепция: ощущение прикосновения и давления на кожу довольно точно локализуется, т.е. относится человеком к определенному участку кожной поверхности. Эта локализация вырабатывается и закрепляется в онтогенезе при участии зрения и проприоцепции. Абсолютная тактильная чувствительность существенно различается в разных частях кожи: от 50 мг до 10 г. Пространственное различение на кожной поверхности, т.е. способность человека раздельно воспринимать прикосновение к двум соседним точкам кожи, также сильно отличается в разных ее участках. На слизистой языка порог пространственного различия равен 0,5 мм, а на коже спины - более 60 мм. Эти отличия связаны главным образом с различными размерами кожных рецептивных полей (от 0,5 мм2 до 3 см2) и со степенью их перекрытия. Температурная рецепция: температура тела человека характеризуется значительным постоянством и поэтому информация о температуре внешней среды, необходимая для деятельности механизмов терморегуляции, имеет особо важное значение. Терморецепторы располагаются в коже, на роговице глаза, в слизистых оболочках, а также в ЦНС - в гипоталамусе. Они делятся на два вида: тепловые (их намного меньше и в коже они лежат глубже) и холодовые. Больше всего терморецепторов в коже лица и шеи. Терморецепторы можно разделить на специфические и неспецифические. Первые возбуждаются лишь температурными воздействиями, вторые отвечают и на механическое раздражение. Большинство терморецепторов имеет локальные рецептивные поля и реагирует повышением частоты генерируемых импульсов, устойчиво длящимся все| время действия стимула. Повышение частоты импульсации происходит пропорционально изменению температуры, причем постоянная частота у тепловых рецепторов наблюдается в диапазоне от 20 до 50 °С, а у холодовых - от 10 до 41°С. Дифференциальная чувствительность терморецепторов велика: достаточно изменить температуру на 0,2°С, чтобы вызвать длительные изменения их импульсации. В определенных условиях холодовые рецепторы могут быть возбуждены и теплом (выше 45 °С). Этим объясняется возникновение острого ощущения холода при быстром погружении в горячую ванну. В настоящее время считают, что наиболее важным фактором, определяющим активность терморецепторов, связанных с ними центральных структур и ощущения человека, является абсолютное значение температуры, а не ее изменения. В то же время интенсивность в начале температурных ощущений зависит от разницы температуры кожи и температуры действующего на нее раздражителя, его площади и места приложения. Кожная механорецепция (механизм возбуждения механорецепторов кожи). Механорецепторы кожи реагируют как на статическое, так и на динамическое воздействие, представляют собой инкапсулированные образования. Механический стимул приводит к деформации мембраны рецептора. В результате этого электрическое сопротивление мембраны уменьшается, т.е. увеличивается ее проницаемость для ионов. Через мембрану рецептора начинает течь ионный ток, приводящий к генерации рецепторного потенциала. При достижении рецепторным потенциалом критического уровня деполяризации генерируются импульсы, распространяющиеся по волокну в ЦНС. Кожная терморецепция (механизм возбуждения терморецепторов кожи). Кожные терморецепторы делятся на холодовые (Реагируют на изменение температуры с коротким латентным периодом. При этом частота ПД линейно зависит от температуры в пределах от 410 до 10°С: чем ниже температура, тем выше частота импульсации) и тепловые (Реагируют на изменение температуры линейно в диапазоне от 20° до 50°С: чем выше температура, тем выше частота генерации ПД. Оптимум чувствительности в пределах 34—43°С). Холодоные подразделяются на специфические - реагирующие только на изменение температуры, и неспецифические - одновременно могут отвечать и на изменение температуры, и на давление. Информация от кожных рецепторов идет в ЦНС по афферентным волокнам группы А-дельта и по волокнам группы С, в ЦНС она доходит с разной скоростью. Импульсация от кожных рецепторов -> спинной мозг, где расположены вторые нейроны -> спиноталамический путь -> вентро-базальные ядра таламуса -> информации поступает в сенсомоторную зону коры больших полушарий и в гипоталамические центры терморегуляции. Мышечно-сухожильная и суставная проприорецепция: Имеется 3 вида рецепторов: мышечные веретена, сухожильные рецепторы Гольджи и суставные рецепторы. Мышечное веретено: одето капсулой и расположено в толще мышцы. Внутри капсулы находится пучок интрафузальных мышечных волокон. Веретена расположены параллельно внешним по отношению к капсуле экстрафузальным волокнам, поэтому при растяжении мышцы нагрузка на веретена увеличивается, а при сокращении – уменьшается. В расслабленной мышце импульсация, идущая от веретен, невелика, но они реагируют повышением частоты разрядов на удлинение мышцы. Таким образом, веретена дают мозгу информацию о длине мышцы и ее изменениях. Импульсация, идущая от веретен, в спинном мозге возбуждает мотонейроны своей мышцы и тормозит мотонейроны мышцы-антагониста, а также возбуждает мотонейроны сгибателей и тормозит мотонейроны разгибателей. Сухожильные рецепторы Гольджи: располагаются в месте перехода мышечных волокон в сухожилия. Отдельный нервный аксон несет афферентные импульсы в спинной мозг и называется аксоном Ib. Он начинается в виде веточек, проходящих между коллагеновыми волокнами сухожилия. Когда мышечные волокна сокращаются, коллагеновые волокна натягиваются и сжимают нервные веточки, которые начинают импульсировать. Таким образом, в результате последовательного крепления сухожильных органов к мышечным волокнам они возбуждаются при укорочении возбужденной мышцы. - суставные рецепторы. Тельца Руффини: находятся в капсуле сустава и воспринимают направление и скорость изменения межзвенного угла. Частота их импульсации возрастает с увеличением скорости изменения суставного угла. Тельца Паччини посылают в ЦНС информацию о положении отдельных частей тела в пространстве и относительно друг друга. Эти рецепторы посылают в ЦНС информацию о значениях межзвенных углов, то есть о положении сустава. Их импульсация продолжается в течение всего периода сохранения межзвенного угла, и она тем больше, чем больше изменения угла. Проводниковый и корковый отделы соматосенсорной системы: - Соматовисцеральная система перерабатывает сенсорную информацию нескольких модальностей, получая ее от кожи, мышц, суставов, внутренних органов и кровеносных сосудов. Она содержит разные типы рецепторов: механорецепторы, терморецепторы, хеморецепторы, осморецепторы, ноцицепторы. Поступающая информация передается по афферентным волокнам в составе чувствительных и смешанных нервов, перерабатывается раздельными проводящими путями и переключательными ядрами, а затем доставляется в соматосенсорную кору, занимающую постцентральные извилины мозга (находящиеся в постцентральной извилине (контрлатеральная поверхность) и области сильвиевой борозды (S-2)) и организованную топически. - Кортикальные клетки, чьи рецептивные поля располагаются на поверхности тела рядом, группируются вместе и в коре. В результате в первичной соматосенсорной коре образуется представительство контрлатеральной поверхности тела, которое может быть картировано с помощью микроэлектродной техники. Карта поверхности туловища и лица человека, находящаяся на постцентралыюй извилине, получила название "сенсорный гомункулус". (Сенсорный гомункулус — это визуализация того, сколько зон мозга активирует сенсорная стимуляция той или иной части тела - чем больше часть, тем больше зон мозга задействуется.) Лемнисковый путь передает в мозг сигналы о прикосновении к коже, давлении на нее и движениях в суставах. Отличительная особенность этого пути – быстрая передача в мозг наиболее точной информации, дифференцированной по силе и месту воздействия. По мере перехода на все более высокие уровни изменяются некоторые важные свойства нейронов лемнискового пути. Значительно увеличиваются рецептивные поля нейронов (в продолговатом мозге – в 2–30, а в коре большого мозга – в 15–100 раз). Несмотря на увеличение размеров рецептивных полей, нейроны остаются достаточно специфичными (нейроны поверхностного прикосновения, глубокого прикосновения, нейроны движения в суставах и нейроны положения или угла сгибания суставов). Для корковой части лемнискового пути характерна четкая топографическая организация, т.е. проекция кожной поверхности осуществляется в центры мозга по принципу «точка в точку» [Кейдель, 1975; Сомьен, 1975; Дудел и др., 1985]. При этом площадь коркового представительства той или иной части тела определяется ее функциональной значимостью: формируется так называемый «сенсорный гомункулюс» (рис. 4.10). Роль соматосенсорной коры состоит в интегральной оценке соматосенсорных сигналов, включении их в сферу сознания и в сенсорное обеспечение выработки новых двигательных навыков. Спиноталамический путь значительно отличается от лемнискового сравнительно медленной передачей афферентных сигналов, нечетко дифференцированной информацией о свойствах раздражителя и не очень четкой ее топографической локализацией; он служит для передачи температурной, всей болевой и в значительной мере тактильной чувствительности. Болевая чувствительность практически не представлена на корковом уровне (раздражение коры не вызывает боли), поэтому считают, что высшим центром болевой чувствительности является таламус, где 60% нейронов в соответствующих ядрах четко реагирует на болевое раздражение. Таким образом, спиноталамическая система играет важную роль в организации генерализованных ответов на действие болевых, температурных и тактильных раздражителей. 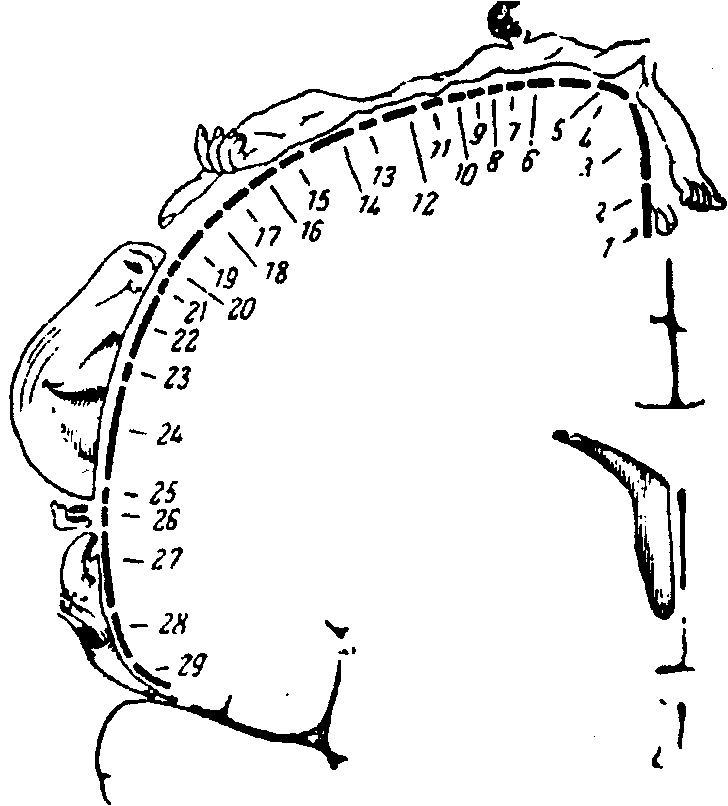 Рис. 4.10. Корковое представительство кожной чувствительности. Расположение в соматосенсорной зоне коры больших полушарий мозга человека проекций различных частей тела: 1– половые органы; 2 – пальцы ноги; 3 – ступня; 4 – голень; 5 – бедро; 6 – туловище; 7– шея; 8 – голова; 9 – плечо; 10–11 – локоть; 12 –предплечье; 13 – запястье; 14 – кисть; 15–19 – пальцы руки; 20 – глаза; 21 – нос; 22 – лицо; 23 – верхняя губа; 24,26 – зубы; 25 – нижняя губа; 27 –язык; 28 – глотка; 29 – внутренние органы. |