Учебное пособие Часть I самара Самарский государственный технический университет 2007
 Скачать 1.81 Mb. Скачать 1.81 Mb.
|

|
В.А. СМИРНОВ, Ю.Н. КЛИМОЧКИН Аминокислоты и полипептиды Учебное пособие Часть I Самара Самарский государственный технический университет 2007  ФЕДЕРАЛЬНОЕ АГЕНТСТВО ПО ОБРАЗОВАНИЮ ФЕДЕРАЛЬНОЕ АГЕНТСТВО ПО ОБРАЗОВАНИЮГОСУДАРСТВЕННОЕ ОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ ВЫСШЕГО ПРОФЕССИОНАЛЬНОГО ОБРАЗОВАНИЯ «Самарский государственный технический университет» ____________________________________________________________ В.А. СМИРНОВ, Ю.Н. КЛИМОЧКИН Аминокислоты и полипептиды Утверждено Редакционно-издательским советом университета в качестве учебного пособия Самара Самарский государственный технический университет 2007 УДК 571.1. + 577.1 С 50 Р е ц е н з е н т канд. мед. наук С. А. Т у м а к о в Смирнов В.А. С 50 Аминокислоты и полипептиды: учеб. пособ. Ч. I./ В.А. Смирнов, Ю.Н. Климочкин. – Самара. Самар. гос. техн. ун-т., 2007. – 110 с.: ил. ISBN 978-5-7964-1057-4 Рассмотрены классификация, номенклатура, строение, физико-химические свойства, методы качественного и количественного анализа аминокислот и полипептидов. Приведены методики качественного и количественного титриметрического и спектрального анализа аминокислот и белков. Предназначено для студентов, обучающихся по специальностям 020101, 260202, 260204, 260401, 240901. УДК 571.1. + 577.1 С 50 ISBN 978-5-7964-1057-4 © Смирнов В.А., Климочкин Ю.Н., 2007 © Самарский государственный технический университет, 2007 ВВЕДЕНИЕ Дисциплина «Биохимия» входит в цикл общепрофессиональных дисциплин для студентов химико-технологического факультета, обучающихся по специальности «Химия» по направлению «Фармацевтическая химия», и факультета пищевых производств Самарского государственного технического университета. За последние 20-25 лет, благодаря успехам молекулярной биологии и широкому внедрению разнообразных современных методов исследования (спектральных, хроматографических и др.), значительно возрос уровень биохимических исследований как в теоретическом, так и в методическом отношении. Исследования проводятся на различных уровнях организации организмов: молекулярном, субклеточном, клеточном, межклеточном, тканевом, органном. В отличие от традиционно изучаемой медиками и биологами дисциплины «Общая биохимия» настоящее учебное пособие по биохимии ставит своей задачей ознакомить студентов с основами и современными аспектами химии живой материи на уровне клетки и не включает в себя биохимию органов и систем организмов. Основное внимание уделено физико-химическим свойствам и методам анализа веществ биологического происхождения. Это обусловлено спецификой деятельности специалистов в области фармацевтической химии и переработки пищевых продуктов. В ассортимент современных лекарственных средств входит значительная группа препаратов биологического происхождения: антибиотики, ферментные препараты, гормоны и др. Эффективная разработка и совершенствование лекарственных средств немыслимы без понимания биохимических процессов в организмах. Все пищевые предприятия перерабатывают сырье биологического происхождения, поэтому курс биохимии является теоретической базой любой пищевой технологии. Целями настоящего учебного пособия являются следующие: 1) ознакомить студентов с биологически активными молекулами природного происхождения, такими, как аминокислоты, белки, ферменты, углеводы, липиды, витамины, гормоны и др., и основными путями обмена веществ на уровне клетки; 2) обучить их основным приемам и навыкам, необходимым для профессионального овладения процессами переработки сырья биологического происхождения, методами оценки его качества и качества препаратов биологического происхождения. 1. ВНЕАУДИТОРНАЯ ПОДГОТОВКА 1.1. АМИНОКИСЛОТЫ 1.1.1. СТРОЕНИЕ, КЛАССИФИКАЦИЯ, НОМЕНКЛАТУРА Аминокислоты – это органические кислоты, содержащие одну или несколько аминогрупп. В зависимости от природы кислотной функции аминокислоты подразделяют на аминокарбоновые, например H2NCH2COOH, аминосульфоновые, например H2N(CH2)2SO2OH, аминофосфоновые и т.д. В живых организмах встречаются, главным образом, аминокарбоновые кислоты (в дальнейшем просто аминокислоты), хотя и редко, но встречаются и аминосульфоновые кислоты, например Н2NCH2CH2SO2OH (таурин – конечный продукт метаболизма цистеина). Классифицируют аминокислоты по разным признакам: по химическому строению, по участию в синтезе белка invivo, по кислотно-основным свойствам, по потребности организмов в данной аминокислоте. По химическому строению аминокарбоновые аминокислоты делятся на аминокислоты ароматического ряда и аминокислоты алифатического ряда. Аминокислоты ароматического ряда составляют лишь весьма небольшой круг веществ, встречающихся в организмах, например пара-аминобензойная кислота (фактор роста микроорганизмов). Основная масса аминокислот, выделенных из разных организмов, относится к классу алифатических аминокислот, т.е. таких аминокислот, в которых амино– и карбоксильная группы связаны с алифатическим атомом углерода. Алифатические аминокислоты делятся в зависимости от относительного положения амино– и карбоксильной групп на α-, β-, γ-аминокислоты и т.д., например: Н2N-CH2-COOH Н2N-CH2-CH2-COOH Н2N-CH2-CH2-CH2-COOH α-Аминоуксусная β-Аминопропионовая γ-Аминомасляная кислота (глицин) кислота (β-аланин) кислота (ГАМК) Основная масса природных аминокислот представлена α-изомерами, а β– и γ-аминокислоты в живых системах представлены единичными случаями. Здесь можно упомянуть нейромедиатор ЦНС млекопитающих γ-аминомасляную кислоту (ГАМК) и β-аланин, являющийся фрагментом кофермента А. α-Аминокислоты чрезвычайно широко распространены в живых организмах любого типа: в растениях, в микроорганизмах, в животных, в морских организмах. Они найдены также в некоторых метеоритах, при этом это такие же аминокислоты, какие входят в состав белков земных организмов. Все α-аминокислоты, кроме глицина, имеют асимметрический (хиральный) α-углеродный атом и существуют в виде двух энантиомеров (L– и D-аминокислот):  L-аминокислота Проекции Фишера D-аминокислота (S-конфигурация) (R-конфигурация) Необходимо отметить, что в случае аминокислот символы L и D приняты для обозначения конфигурации α-углеродного атома, а не знака оптического вращения (для указания последнего используются символы + и −). Например, L-цистеин в нейтральном водном растворе является левовращающим (−), напротив, L-аланин – правовращающим (+), хотя также принадлежит к L-ряду. На знак оптического вращения оказывает влияние среда, в которой проводится определение. Так, L-серин в нейтральной среде является лево-, а в кислой – правовращающим. А 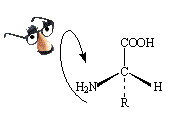 бсолютные конфигурации устанавливаются с помощью правила последовательности Куна, Ингольда, Прелога. Для определения конфигурации асимметрического центра Сabdf сначала определяют старшинство групп abdf. Чем больше атомный номер первого атома группы, тем она старше; если они одинаковы, сравнивают атомные номера вторых атомов группы и т.д. Так, в случае α-аминокислот последовательность, соответствующая уменьшению старшинства групп, такова: H2N, COOH, R, H. бсолютные конфигурации устанавливаются с помощью правила последовательности Куна, Ингольда, Прелога. Для определения конфигурации асимметрического центра Сabdf сначала определяют старшинство групп abdf. Чем больше атомный номер первого атома группы, тем она старше; если они одинаковы, сравнивают атомные номера вторых атомов группы и т.д. Так, в случае α-аминокислот последовательность, соответствующая уменьшению старшинства групп, такова: H2N, COOH, R, H.Д Все протеиногенные аминокислоты (определение термина см. далее) относятся к L-ряду. Однако за последнее время появилось много данных о наличии D-аминокислот как в свободном состоянии, так и в составе пептидов, главным образом среди продуктов метаболизма микроорганизмов. Остатки D-аминокислот входят в состав многих антибиотиков, например, полимиксин D содержит фрагмент D-серина, грамицидин А – D-лейцина, актиномицин С – D-валина. Некоторые природные α-аминокислоты (изолейцин, треонин) имеют два асимметрических атома углерода и соответственно существуют в виде двух пар диастереомеров. Эти аминокислоты, встречающиеся в природных белках, относят к нормальному ряду, а соответсвующие им стереоизомеры, полученные синтетическим путем, обозначают как алло-аминокислоты:  L-треонин D-треонин L-алло-треонин D-алло-треонин 2-(S), 3-(R) 2-(R), 3-(S) 2-(S), 3-(S) 2-(R), 3-(R)  Симметричная аминокислота цистин может существовать в трех формах: L-, D- и внутренне компенсированной оптически неактивной – мезо-форме. Природный L-цистин образуется в организме при окислении цистеина, что играет большую роль в формировании нативной структуры белка при его посттрансляционном созревании:  L-цистин D-цистин мезо-цистин По участию в синтезе белка invivo аминокислоты делятся на протеиногенные и непротеиногенные. Протеиногенными называют аминокислоты, которые включаются в состав белков в ходе биосинтеза в рибосомах под генетическим контролем информационной РНК. Их ровно 20, и все они относятся к L-ряду (табл.1.1). В то же время из природных белков выделено к настоящему времени около 30 аминокислот, например L-оксипролин, L-цистин, L-β-оксиглутаминовая кислота и др.:  L-Оксипролин L-β-оксиглутаминовая кислота 4(R)-гидрокси-2(S)-пролин 3(R)-гидрокси-2(S)-глутаминовая кислота Этот факт объясняется тем, что линейная немодифицированная полипептидная цепь превращается в полноценный функциональный белок в результате многостадийного процесса (созревание), который начинается сразу же после начала трансляции. В процессе созревания белка осуществляются различные ферментативные реакции, такие, как окисление, фосфорилирование, N-гликозилирование, взаимодействие с белками связывания (шапероны – белки, направляющие процесс свертывания цепи). В результате процесса созревания образуются характерные для данного белка вторичная и третичная структуры. Н  апример, предшественник коллагена – линейный проколлаген – прежде чем превратиться в коллаген, вступает в ряд ферментативных реакций. Он подвергается окислению, которое заключается в гидроксилировании остатков пролина и лизина, а также в образовании дисульфидных мостиков между остатками цистеина. Эти процессы обеспечивают правильное скручивание полипептидных цепей в тройную спираль коллагена. апример, предшественник коллагена – линейный проколлаген – прежде чем превратиться в коллаген, вступает в ряд ферментативных реакций. Он подвергается окислению, которое заключается в гидроксилировании остатков пролина и лизина, а также в образовании дисульфидных мостиков между остатками цистеина. Эти процессы обеспечивают правильное скручивание полипептидных цепей в тройную спираль коллагена.Таблица 1.1 – Протеиногенные аминокислоты
Коферментом, участвующим в процессе окисления остатков L-пролина в проколлагене, является аскорбиновая кислота, дефицит которой вызывает торможение процесса образования коллагена из проколлагена (заболевание – цинга, рис.1.1).  Р и с.1.1. Гидроксилирование остатка пролина в проколлагене: 1 – фермент – проколлагенпролин-4-диоксигеназа-[аскорбат]; 2 – остаток L-пролина; 3 – остаток -4-гидрокси-L-пролина (оксипролина); 4 – аскорбиновая кислота; 5 – дегидроаскорбиновая кислота Кроме протеиногенных и образующихся из них аминокислот, входящих в состав белка, но не являющихся протеиногенными, в организмах обнаружено большое количество непротеиногенных аминокислот, которые не входят в состав белков. Только в растениях и микроорганизмах обнаружено более 200 аминокислот алифатического ряда. Приведем лишь несколько примеров, иллюстрирующих большое разнообразие непротеиногенных аминокислот (рис.1.2). Среди них обнаружены соединения и с различными циклами, в том числе гетероциклами, и с непредельными связями, и с такими элементами, как хлор, бром, йод, селен, и аналоги протеиногенных аминокислот с D-конфигурацией. Что касается D-энантиомеров протеиногенных аминокислот, таких, как D-аланин, D-валин, D-цистеин, D-фенилаланин, D-глутамин и другие, то такие аминокислоты часто входят в состав полипептидных антибиотиков, продуцируемых микроорганизмами, и являются компонентами клеточных стенок многих бактерий. |

